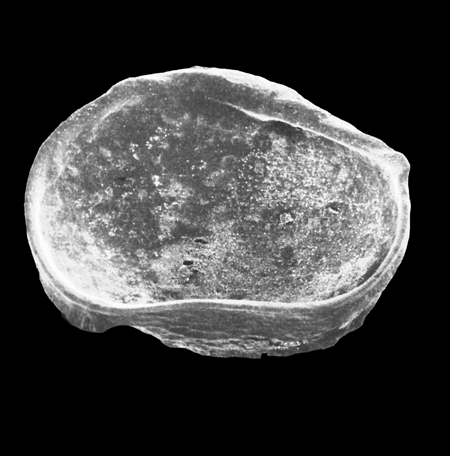
| Introduction |
| Ostracods are by far the most complex organisms studied within the field of micropalaeontology. They are Metazoa and belong to the Phylum Arthropoda (as trilobites), Class Crustacea (as lobsters and crabs). An important distinguishing feature Ostracods share with other arthropods is the bilateral symmetry of their body form. The paired body parts are enclosed in a dorsally hinged carapace composed of low magnesium calcite, which is what is commonly preserved in the fossil record. They are found today in almost all aquatic environments including hot springs, caves, within the water table, semi-terrestrial environments, in both fresh and marine waters, within the water column as well as on (and in) the substrate. In fact almost anywhere that's wet, even if only for a brief period! |
 |
| History of Study |
| The oldest generic names given to ostracods are Cypris and Cythere by Muller in the 1770's and 80's, these are now commonly used as suffixes and prefixes in ostracod nomenclature. In the 1860's Sars classified ostracods as an order divided into four suborders, Podocopa, Myodocopa, Cladocopa and Platycopa. In the early 20th Century workers in the Appalachians in the U.S.A classified Palaeozoic ostracods. In 1958 Pokorny combined these two classifications and in 1961 an Anglo American treatise modified Pokorny's work to give the foundation of today's classification system. It was not until Pokorny's work that the fossil and living classifications were linked. |
|
| Range |
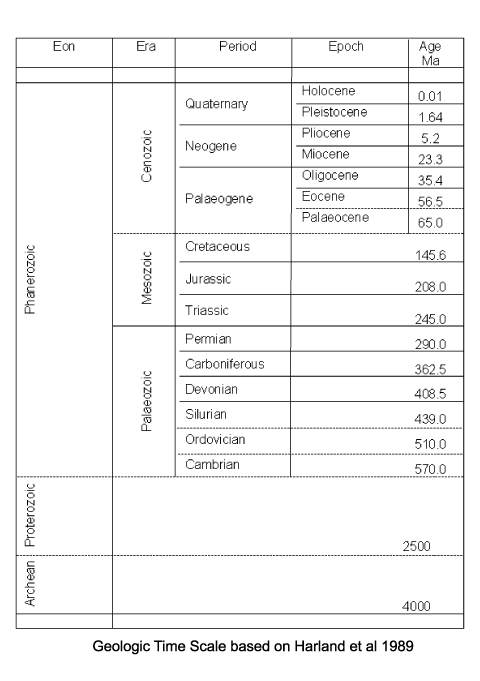
| Ostracod-like organisms (bivalved arthropods) are recorded from the Cambrian, but it is uncertain whether these can be classified as true ostracods. Myodocopid and podocopid forms are recorded from the Ordovician. All these early forms are marine, the first freshwater forms (Darwinulacea and Carbonita) occur in the Carboniferous and by the Jurassic ostracods are common in freshwater environments. Between the Silurian/Devonian and the present there are big gaps in the fossil record of planktonic marine forms, which is thought to reflect weak calcification of the carapace. |
 |
|
| Classification |
| The Class Ostracoda is separated from other Crustacea by their laterally compressed body, undifferentiated head, seven or less thoracic limbs and the bivalved, perforate carapace lacking growth lines. The living ostracods are further classified in many cases by variations in their appendages and other soft parts. Although exceptionally well preserved fossil ostracods with the soft parts intact have been found these are very rare and therefore the morphological features (see below) of the carapace have become vital in fossil ostracod classification. The Ostracoda have been divided into five Orders, the extant Podocopida and Myodocopida and the extinct Phosphatocopida, Leperditicopida and Palaeocopida (however, the latter groups may well not be ostracods in the strict biological sense). |
|
| Applications |
| Since the fossil record of planktonic marine ostracods is so patchy, biostratigraphic uses of ostracods based on benthic forms are limited to specialised marine environments for example in the Jurassic of the North Sea. In the marine environment benthic ostracods are utilised for palaeoenvironmental reconstructions. Freshwater and brackish facies commonly contain abundant ostracods which are used for environmental studies and for biostratigraphic zonations, for instance in non-marine sediments from Mongolia and China. |
|
| Biology |
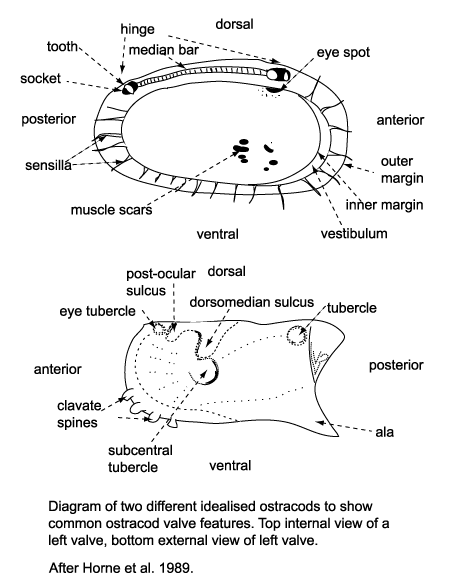 |
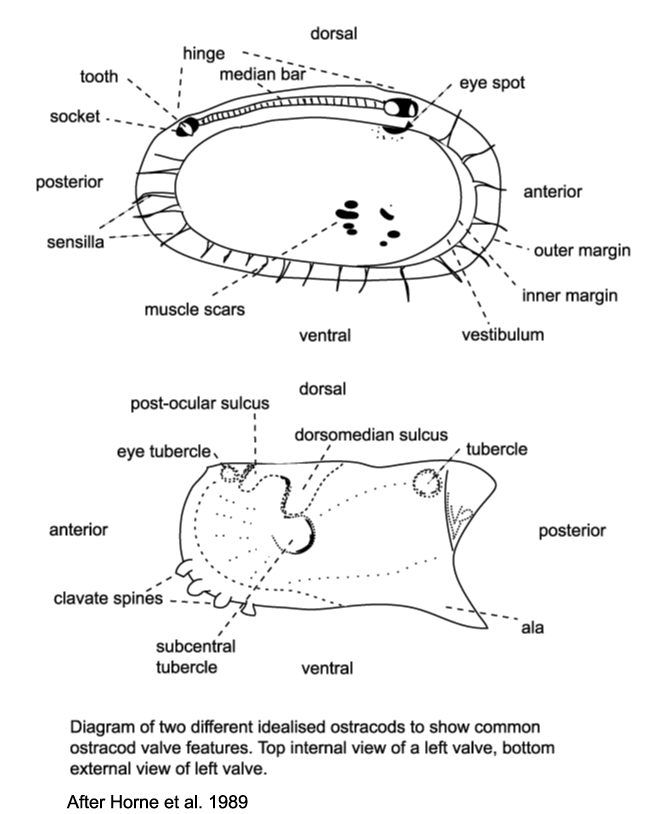
Several morphological features of ostracods are at times preserved
in the fossil forms and have been utilised in their classification. The ostracod carapace is usually ovate, kidney-shaped or bean-shaped, it is divided into a right and left valve, one being, commonly slightly larger than the other partially overlapping it, and hinged at the dorsal margin. The hinge is an important feature in terms of taxonomy and classification. Four basic types of hinge are recognised:
- The adont hinge is the simplest, without teeth or sockets, often forms part of a contact groove on the larger valve and a corresponding ridge on the smaller valve.
- The merodont hinge is composed of a tooth and socket at each end of a groove or ridge structure (complementary negative and positive structures in left and right valves).
- The entomodont hinge differs from the merodont hinge style by having a coarsely crenulated anterior portion of the median groove/ ridge element.
- The amphidont hinge has a more complex median structure with an anterior tooth and socket.
|
| When alive the two valves of the carapace are closed by adductor muscles, these are normally connected to the inner surface of the valve at a point just anterior of the valve centre, they frequently leave scars on the valves inner surface and either a subcentral tubercle (a sort of boss) or a sulcus (an elongate shallow depression) on the outside. The podocopid ostracods produce a calcified overlapping flange around the ventral margin called a duplicature. |
| Ostracods sense their surroundings using sensilla (hairs or bristles) which project through the carapace via pore canals, at the margins these are called marginal pore canals. Some shallow water ostracods have eyes and their carapaces have clear eyespots or raised eye tubercles. |
| Ostracods can reproduce sexually and asexually (parthenogenesis).
Ostracods show sexual dimorphism, that is males and females of the same species have carapaces of differing size and shape.They grow by moulting up to nine growth stages (instars). Because of this sexual dimorphism and the ontogenetic variations of ostracods great care must be taken with taxonomy, as a single species may have a series of juvenile stages as well as two adult morphotypes. |
| The ecology of ostracods is often reflected in the shape and structure of their carapaces hence making them useful palaeoenvironmental indicators. Freshwater ostracods in general tend to have smooth, thin, weakly calcified simple bean-shaped carapaces. They feed on a wide range of food stuffs including diatoms, bacteria and detritus. Pelagic ostracods also tend to have thin, smooth shells and may have long powerful swimming appendages or antennules which have led to the formation of rostral incisures at the anterior of the carapace to allow freer movement of these appendages. Benthic ostracods are commonly detritivores or filter feeders, they either burrow into the substrate, in which case their carapaces tend to be smooth, small, robust and sometimes elongated. Epifaunal types may have flattened ventral surfaces sometimes with projecting alar wings, frills, keels or lateral spines. Those found on coarser substrates in higher energy environments tend to have more robust heavily ribbed or reticulated carapaces. |
|
| Life Cycle |
| Ostracods like other Crustacea moult between growth stages (called an instar), this process is known as ecdysis. There are usually nine instars between egg and adult. This fact has extremely important implications for palaeontological studies. For example, if an assemblage contains a mix of instars it is relatively safe to assume the material is in situ (a biocoenosis, a true reflection of the living assemblage). Ostracods also have a variety of complex reproductive strategies, including brood care of eggs within the carapace (e.g. Darwinula), desiccation-resistant eggs (known to survive in a dry state only to hatch on immersion in water years later), sexual and asexual strategies including parthenogenesis which in Darwinula is thought to be the only method of reproduction utilised. Parthenogenesis is asexual reproduction via the female only, there appears to have been no male Darwinula's for many thousands if not millions of years! |
|
| Preparation Technique |
| WARNING:
Please remember all preparation techniques require the use of hazardous materials and equipment and should only be caried out in properly equiped laboratories, wearing the correct safety clothing and under the supervision of qualified staff. |
| Ostracod carapaces range in size from approximately 100 microns up to several millimetres, and they are commonly prepared in the same way as foraminifera with careful washing with hydrogen peroxide and/or washing soda and sieving using a standard 63 micron sieve. Several washes may be required to break down well indurated material and care should be taken when washing through the sieve to prevent breakage of the specimens. The cleaned residue can then be dried, sieved into fractions (generally 63-125 microns, 125-250 microns, 250-500 microns and greater than 500 microns) and "picked". Care must be taken to clean all sieves and materials used between the preparation of each sample to prevent contamination. |
|
| Observation Techniques |
| Once fossil samples have been prepared ostracod carapaces can be picked from any remaining sediment using a fine brush and a reflected light, binocular microscope. The best method is to scatter a fine dusting of sieved sediment on to a black tray divided into squares, this can then be scanned under the microscope and any ostracods preserved in the sediment can be picked out with a fine brush (preferably a 000 sable paint brush). The picked specimens can then be mounted in card slides divided into numbered squares with sliding glass covers. Gum tragocanth was traditionally used to attach the specimens to the slides but modern office type water soluble paper adhesives are now used. Ostracods are large enough to be observed live in wet preps under microscopes and sometimes with the naked eye. Almost any relatively still water will contain ostracods and samples can be collected especially by scraping them from the surface of water plants or sediment. |
|
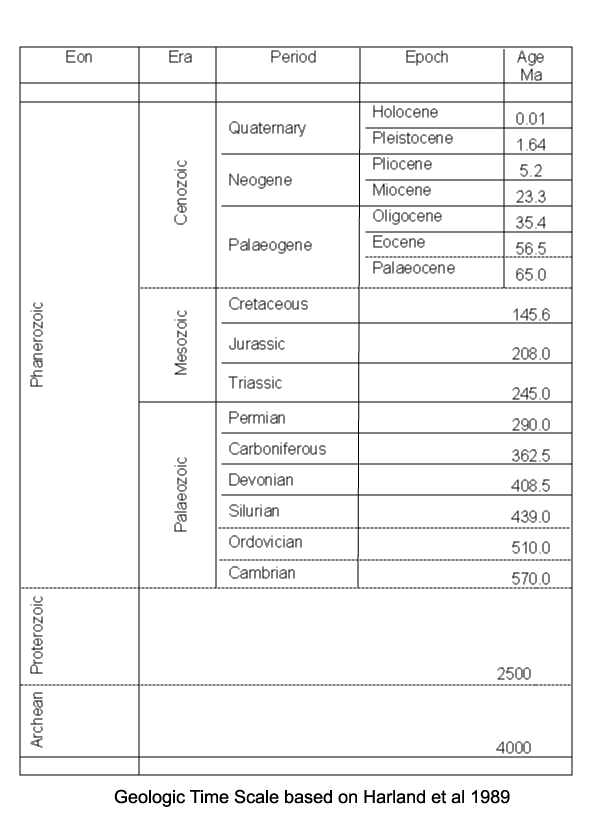
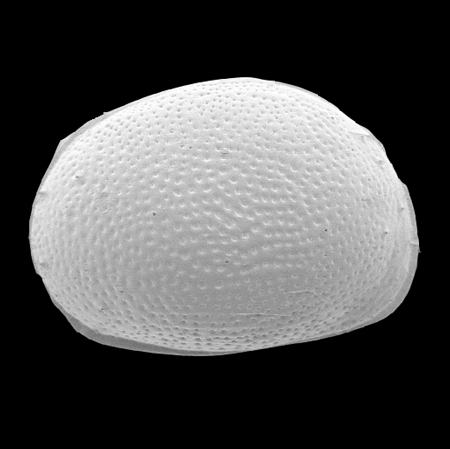
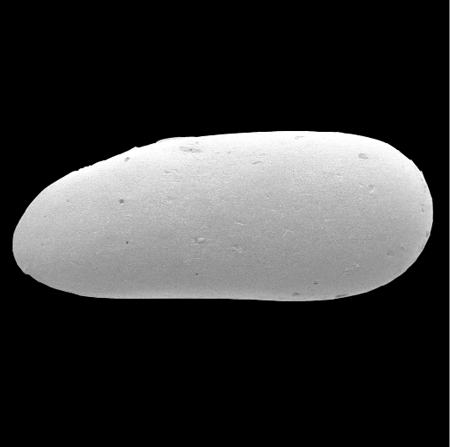
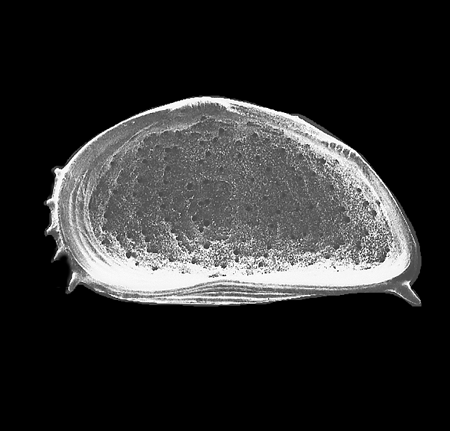
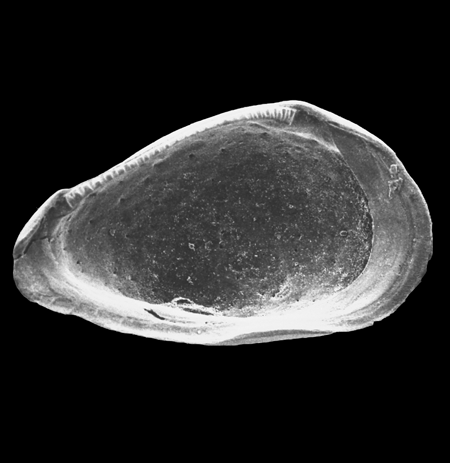
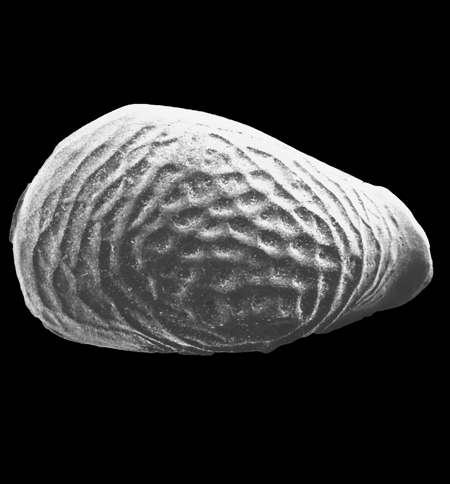
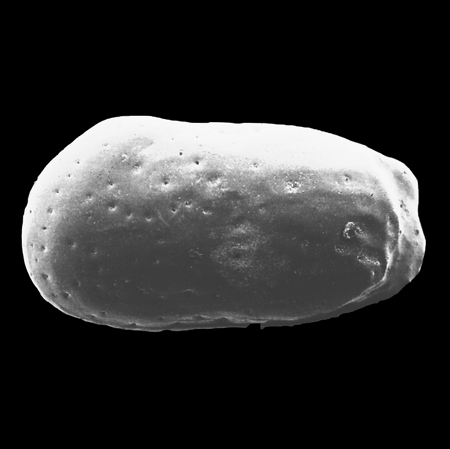
| Images |
| The following images are of a representative selection of ostracods aimed at giving a general overview of the different morphotypes. Each specimen is given a generic and, if possible, a species name followed by its age range, the site location from which the sample was obtained and its size in microns. LM (Light Microscope) SEM (Scanning Electron Microscope). Typical and selected marker species are illustrated from each main period of the geological column in which ostracods occur. Click on an image to view a larger version. Because Ostracods are such a diverse taxa they have been split into two groups: freshwater and marine forms. |
|
|
| Freshwater |

|
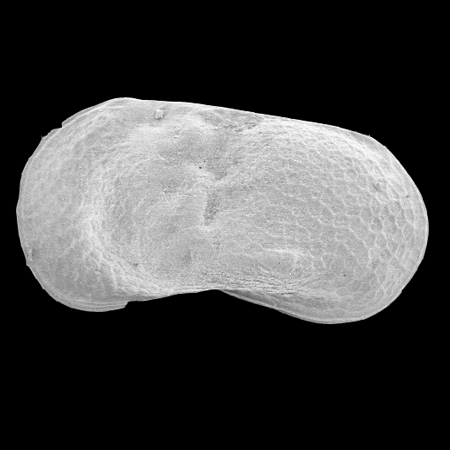
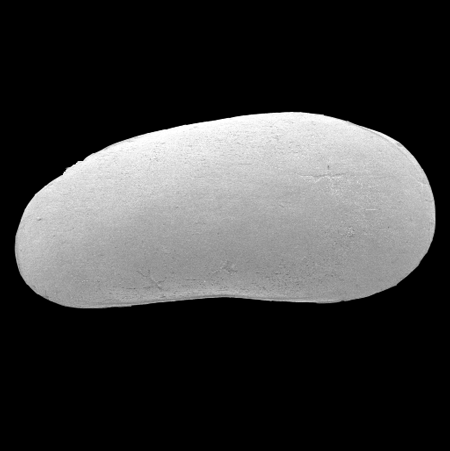
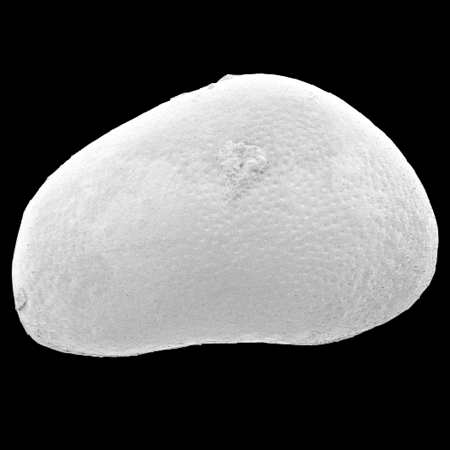
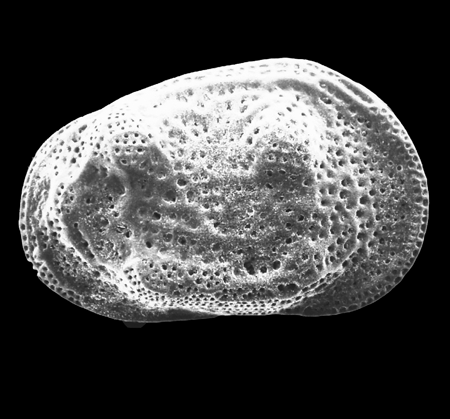
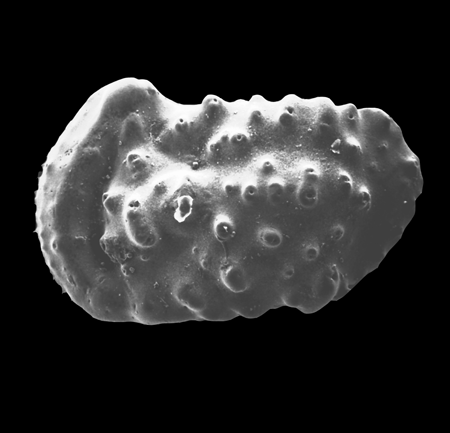
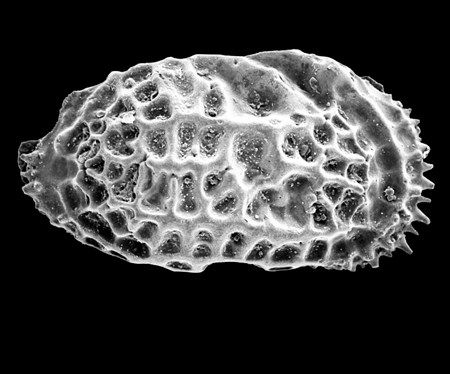
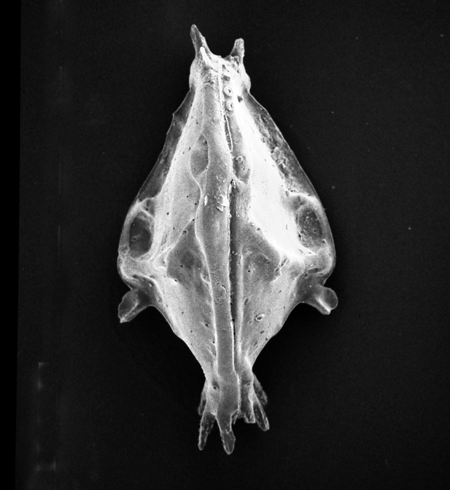
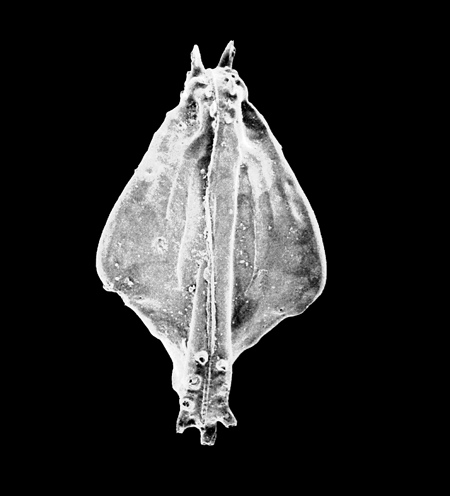
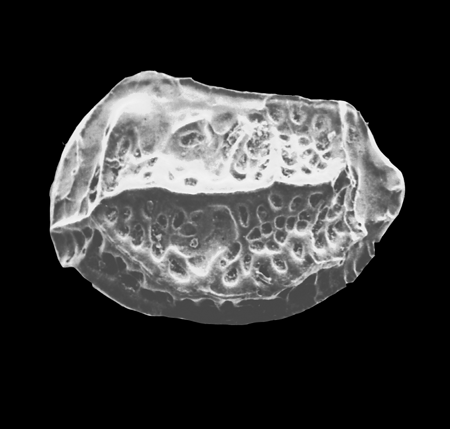
| Potamocypris
sp.
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| ~600 microns right valve SEM |

|
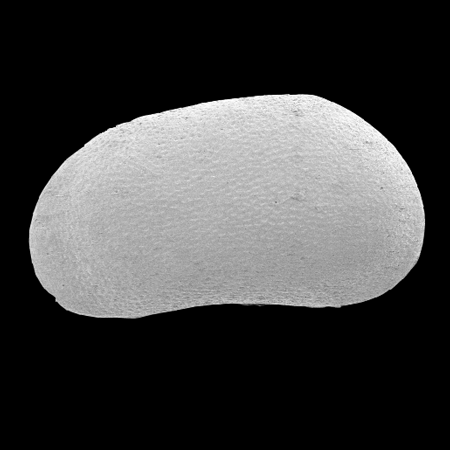
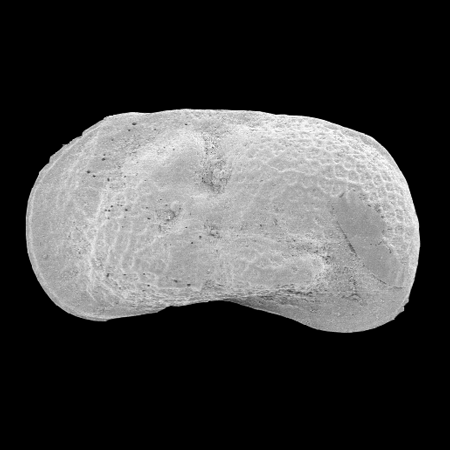
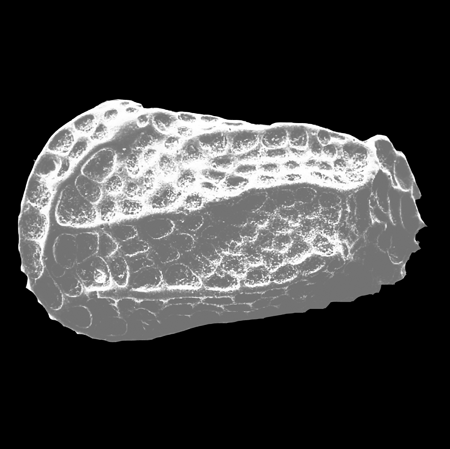
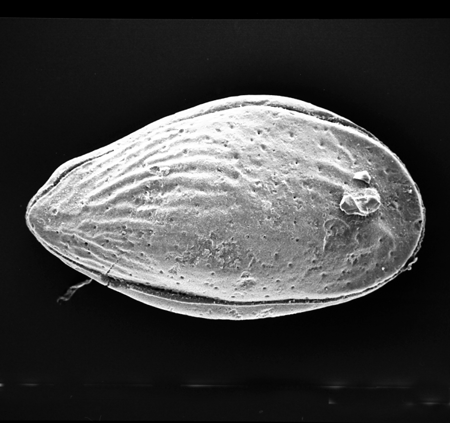
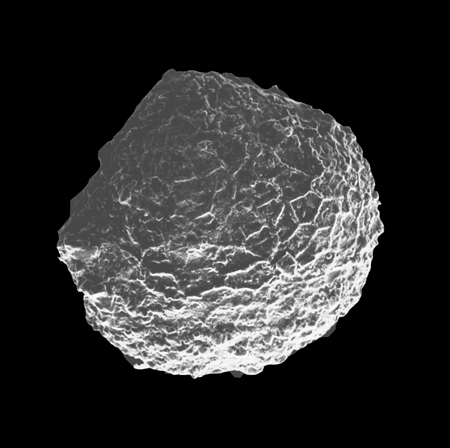
| Cyclocypris
sp.
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| ~500 microns left valve SEM |

|
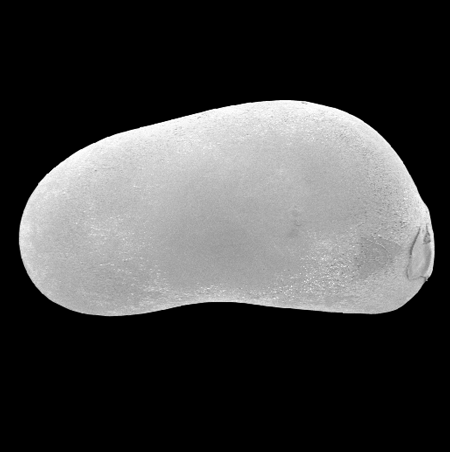
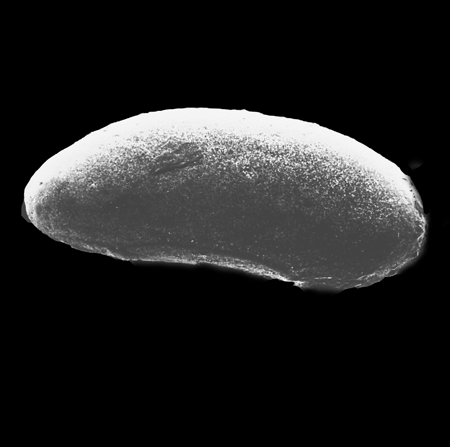
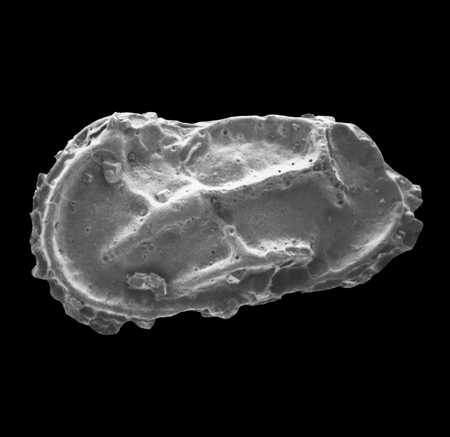
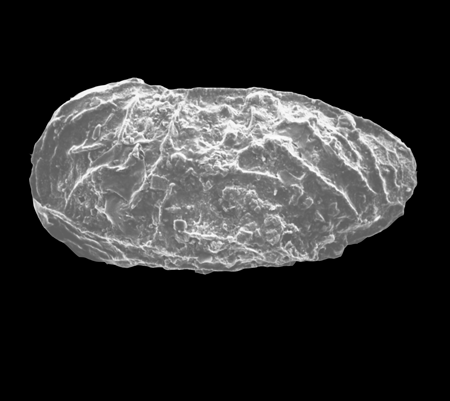
| Cypria ophtalmica
(Jurine)
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 650 microns left valve SEM |
|
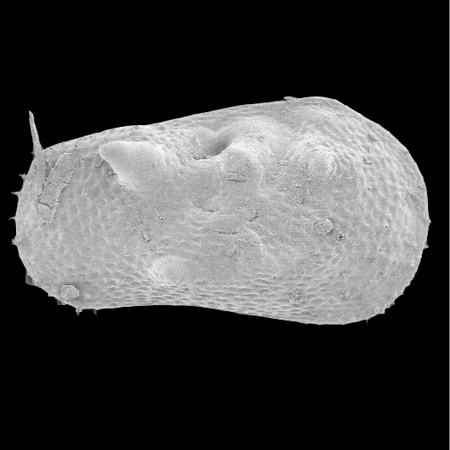
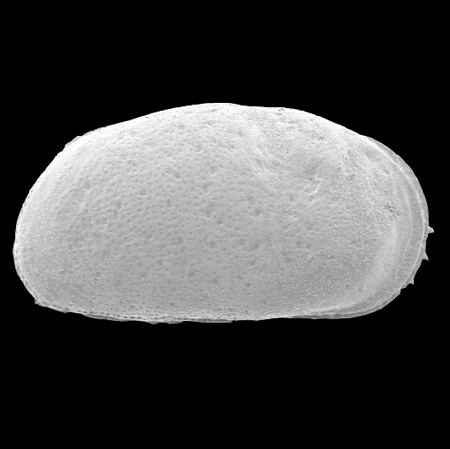
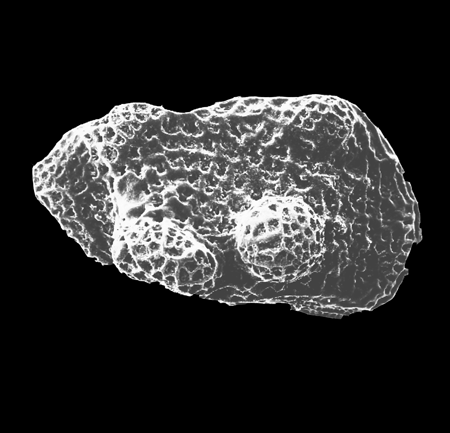
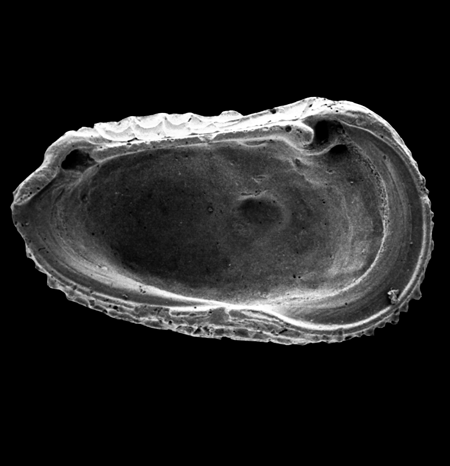
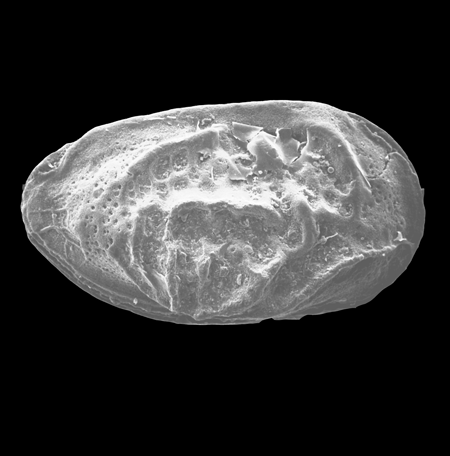
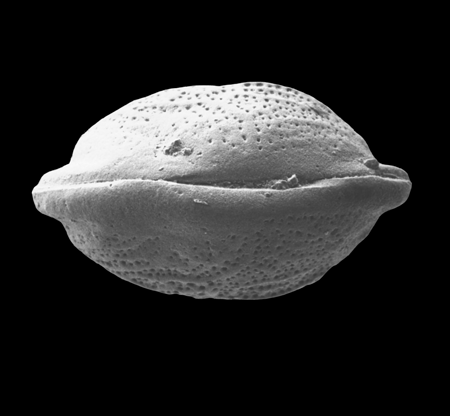
| Ilyocypris
sp.
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 800 microns right valve SEM |

|
| Metacypris cordata
Brady and Robertson
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 520 microns right valve SEM |

|
| Darwinula stevensoni
Brady and Robertson
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 780 microns right valve SEM |

|
| Cyclocypris ovum
(Jurine)
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 480 microns right valve SEM |

|
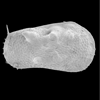
| Limnocythere sanctipatricii
(Brady and Robertson)
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 850 microns female left valve SEM |
|
| Ilyocypris
sp.
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 750 microns right valve SEM |

|
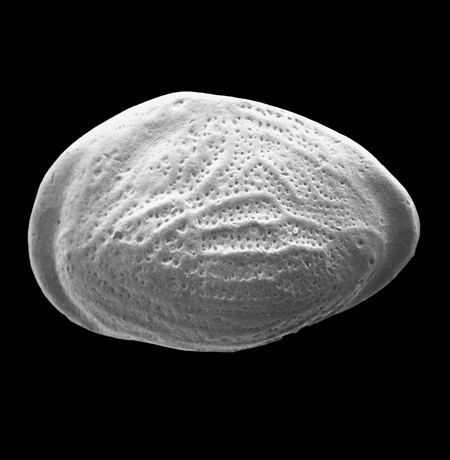
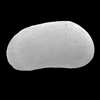
| Cyclocypris laevis
Muller
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 580 microns dorsal view carapace SEM |
|
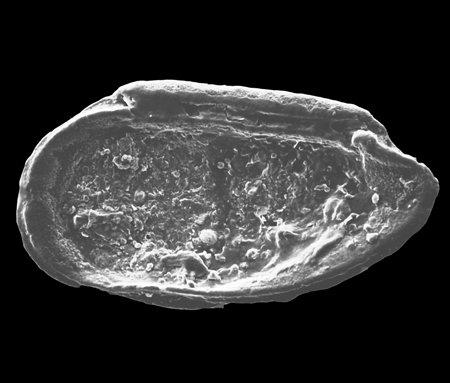
| Pseudocandona rostrata
(Brady and Norman)
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 1000 microns left valve SEM |

|
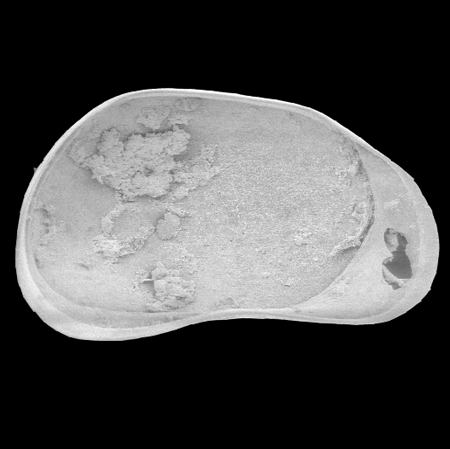
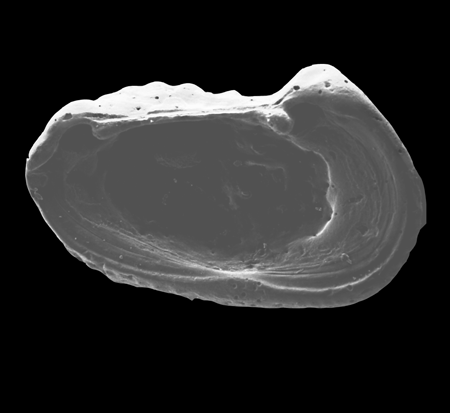
| Pseudocandona rostrata
(Brady and Norman)
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 1000 microns left valve internal view SEM |

|
| Candona candida
(Muller)
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 1100 microns female right valve SEM |

|
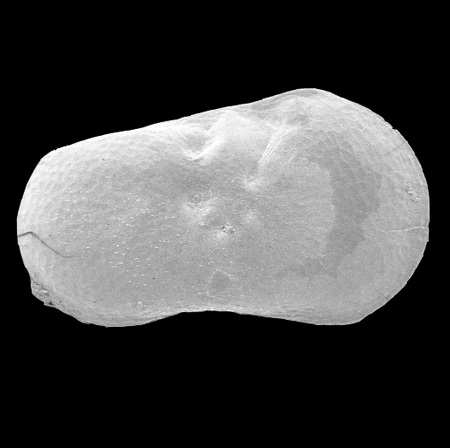
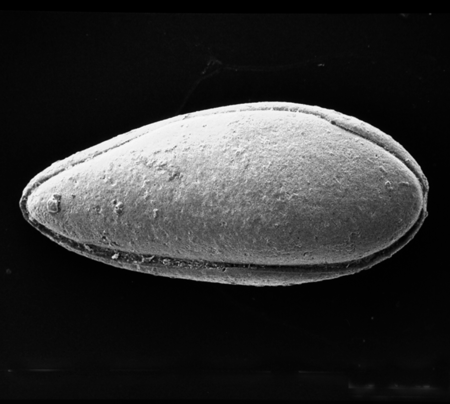
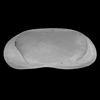
| Herpetocypris
sp.
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 2000 microns female right valve SEM |
|
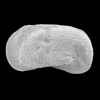
| Herpetocypris reptans
(Baird)
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 2000 microns internal view right valve SEM |
|
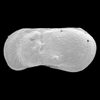
| Limnocythere inopinata
(Baird)
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 630 microns female left valve SEM |
|
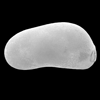
| Limnocythere sanctipatricii
(Brady and Robertson)
|
| ?-Recent
|
| Mooghaun Lough, County Clare, Eire |
| 840 microns female left valve SEM |
|
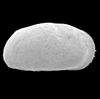
| Candona neglecta
(Sars)
|
| ?-Recent
|
| Merja Sidi Bou Rhaba, Northwestern Morocco |
| 1300 microns SEM |
|
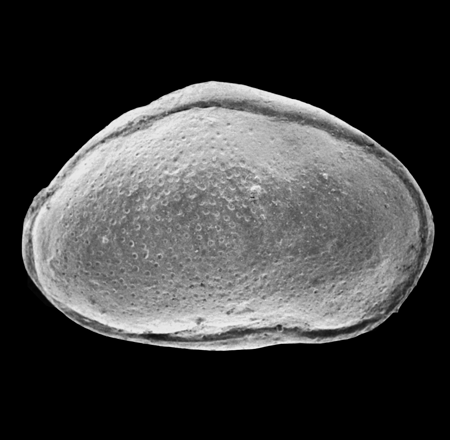
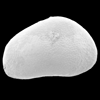
| Cyprideis torosa
(Jones)
|
| ?-Recent
|
| Merja Sidi Bou Rhaba, Northwestern Morocco |
| SEM |

|
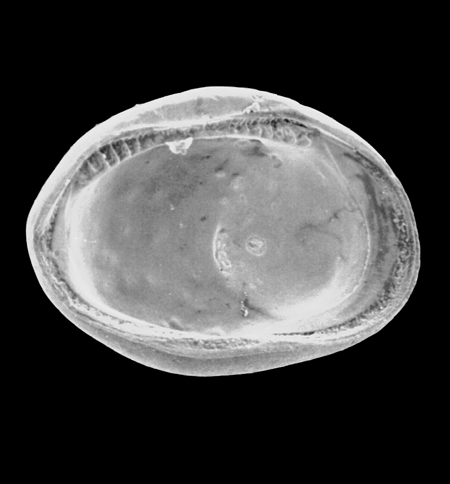
| Cyprideis torosa
(Jones) detail of sieve pores with setae
|
| ?-Recent
|
| Merja Sidi Bou Rhaba, Northwestern Morocco |
| SEM |
|
| Potamocypris
sp. (Brady)
|
| ?-Recent
|
| Merja Sidi Bou Rhaba, Northwestern Morocco |
| SEM |

|
| Ilyocypris gibba
(Ramdohr)
|
| ?-Recent
|
| Merja Sidi Bou Rhaba, Northwestern Morocco |
| 820 microns SEM |
|
| Marine |

|
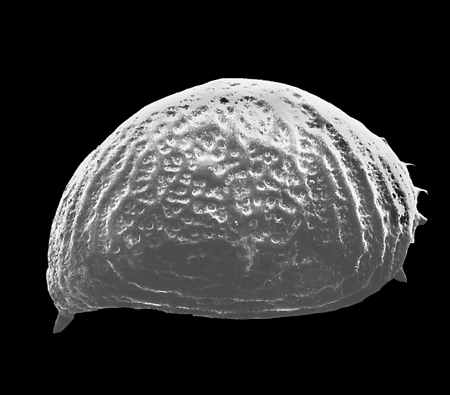
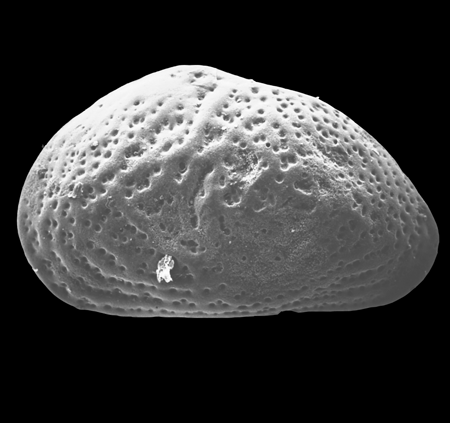
| Heterocyprideis sorbyana
(Jones)
|
| ?-Recent
|
| Skagen Core, Jutland, Denmark |
| adult right valve SEM |

|
| Heterocyprideis sorbyana
(Jones)
|
| ?-Recent
|
| Skagen Core, Jutland, Denmark |
| adult right valve internal view SEM |

|
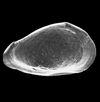
| Rabilimis mirabilis
Brady
|
| ?-Recent
|
| Skagen Core, Jutland, Denmark |
| adult left valve SEM |
|
| Cytheropteron pseudomontrosiense
Whatley and Masson
|
| ?-Recent
|
| Skagen Core, Jutland, Denmark |
| adult left valve internal view SEM |

|
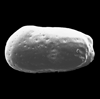
| Cytheropteron pseudomontrosiense
Whatley and Masson
|
| ?-Recent
|
| Skagen Core, Jutland, Denmark |
| juvenile left valve SEM |
|
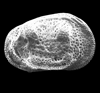
| Normanicythere leioderma
(Norman)
|
| ?-Recent
|
| Skagen Core, Jutland, Denmark |
| adult left valve SEM |
|
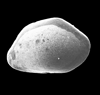
| Cluthia cluthae
(Brady, Crosskey and Robertson)
|
| ?-Recent
|
| Skagen Core, Jutland, Denmark |
| adult right valve SEM |
|
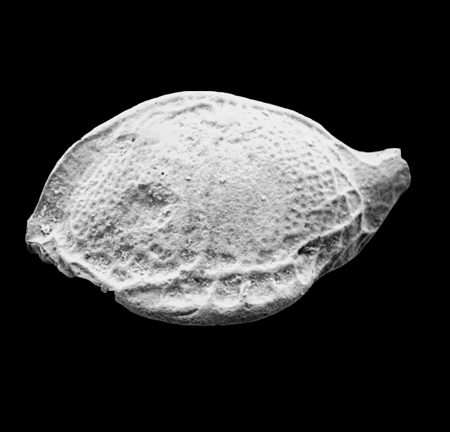
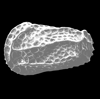
| Bairdia subdeltoidea
(von Munster)
|
| Lower Miocene-Pliocene
|
| Pissouri, Cyprus, Eastern Mediterranean |
| adult right valve SEM |
|
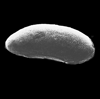
| Cistacythereis pokornyi hellenica
Uliczny
|
| Pliocene-Recent
|
| Pissouri, Cyprus, Eastern Mediterranean |
| adult left valve SEM |
|
| Australoecia poteroacuta
Coles and Whately
|
| Upper Eocene-Upper Oligocene
|
| Pissouri, Cyprus, Eastern Mediterranean |
| right valve SEM |

|
| Eucytherura complexa
(Brady)
|
| Pliocene-Recent
|
| Pissouri, Cyprus, Eastern Mediterranean |
| right valve SEM |

|
| Aurila punctata punctata
(von Munster)
|
| Late Miocene-Recent
|
| Pissouri, Cyprus, Eastern Mediterranean |
| left valve SEM |

|
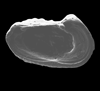
| Cythere texana
Stadnichenko
|
| ?Eocene?
|
| Pin Oak Creek, Bastrop County, Texas, USA |
| left valve (femle) SEM |
|
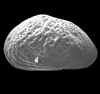
| Cythere texana
Stadnichenko
|
| ?Eocene?
|
| Pin Oak Creek, Bastrop County, Texas, USA |
| left valve internal view (male) SEM |
|
| Cytheridea (Clithrocytheridea) cf. oliveri
(Cushman)
|
| ?Eocene?
|
| Pin Oak Creek, Bastrop County, Texas, USA |
| left valve (female) SEM |
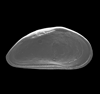
|
| Cytheridea (Clithrocytheridea) cf. oliveri
(Cushman)
|
| ?Eocene?
|
| Pin Oak Creek, Bastrop County, Texas, USA |
| left valve internal view (female) SEM |

|
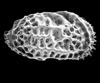
| Semicytherura
Sp.
|
| Late Palaeocene
|
| Itori Borehole, Eastern Dahomey Basin, Nigeria |
| 560 microns (length) SEM |
|
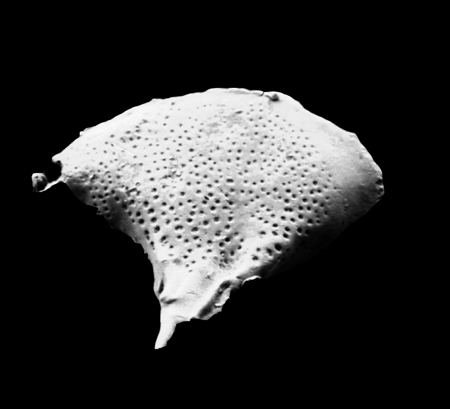
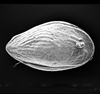
| Paleocosta olurebei
(Reyment)
|
| Late Palaeocene
|
| Araromi Borehole, Eastern Dahomey Basin, Nigeria |
| 760 microns (length) SEM |
|
| Buntonia ioruba
Reyment
|
| Late Maastrchtian-Late Palaeocene
|
| Araromi Borehole, Eastern Dahomey Basin, Nigeria |
| 590 microns (length) right valve female SEM |

|
| Stigmatocythere teiskotensis
(Apostolescu)
|
| Palaeocene
|
| Araromi Borehole, Eastern Dahomey Basin, Nigeria |
| 810 microns (length) ventral view female SEM |

|
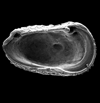
| Costa levigata
|
| Palaeocene
|
| Itori Borehole, Eastern Dahomey Basin, Nigeria |
| 780 microns (length) left valve female SEM |
|
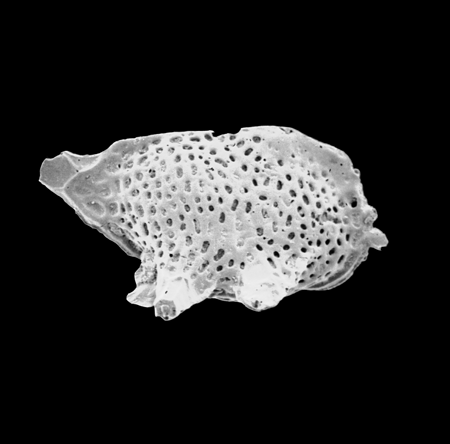
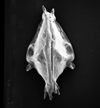
| Actinocythereis asanmamoi
Reyment
|
| Late Palaeocene
|
| Gebekebo Borehole, Eastern Dahomey Basin, Nigeria |
| 660 microns (length) left valve internal view female SEM |
|
| Brachycythere armata
Reyment
|
| Late Maastrichtian (latest Cretaceous)
|
| Araromi Borehole, Eastern Dahomey Basin, Nigeria |
| 810 microns (length) dorsal view SEM |

|
| Brachycythere armata
Reyment
|
| Late Maastrichtian (latest Cretaceous)
|
| Araromi Borehole, Eastern Dahomey Basin, Nigeria |
| 810 microns (length) ventral view SEM |

|
| Parascypris
sp.
|
| Late Maastrichtian (latest Cretaceous)
|
| Araromi Borehole, Eastern Dahomey Basin, Nigeria |
| 960 microns (length) ventral view SEM |

|
| Phacorhabdotus pokornyi
Paulson
|
| Late Cretaceous-Palaeocene
|
| Eastern Gulf Coast, U.S.A |
| 580 microns (length) left valve SEM |

|
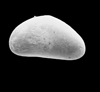
| Cytherelloidea crafti
Sexton
|
| Late Cretaceous?
|
| Eastern Gulf Coast, U.S.A |
| 700 microns (length) left valve SEM |
|
| Bythocypris windhami
Butler and Jones
|
| Early Cretaceous-Recent?
|
| Eastern Gulf Coast, U.S.A |
| 920 microns (length) left valve SEM |

|
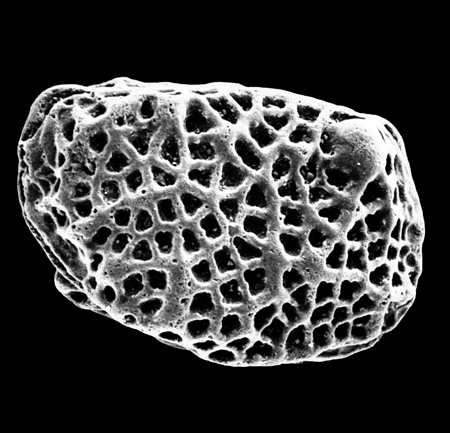
| Carinocythereis priddyi
Smith
|
| Cretaceous?
|
| Eastern Gulf Coast, U.S.A |
| 670 microns (length) left valve SEM |

|
| Cythereis dallasensis
Alexander
|
| Cretaceous?
|
| Eastern Gulf Coast, U.S.A |
| 840 microns (length) left valve SEM |

|
| Antibythocypris bipunctata
|
| Maastrichtian?
|
| Eastern Gulf Coast, U.S.A |
| 360 microns (length) left valve SEM |

|
| Antibythocypris bipunctata
|
| Maastrichtian?
|
| Eastern Gulf Coast, U.S.A |
| 360 microns (length) internal view right valve SEM |

|
| Cytheropteron gaudaloupense
Crane
|
| Cretaceous-Recent?
|
| Eastern Gulf Coast, U.S.A |
| 660 microns (length) left valve SEM |

|
| Cytheropteron furcalatum
Alexander
|
| Cretaceous-Recent?
|
| Eastern Gulf Coast, U.S.A |
| 680 microns (length) SEM |

|
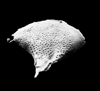
| Brachycythere ledoforma porosa
Crane
|
| Cretaceous-Recent?
|
| Eastern Gulf Coast, U.S.A |
| 680 microns (length) SEM |
|
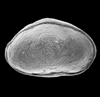
| Cytheropteron harrissi
Skinner
|
| Cretaceous-Recent?
|
| Eastern Gulf Coast, U.S.A |
| 690 microns (length) SEM |
|
| Ovocytheridea reniformis
van den Bold
|
| Cenomanian-Turonian (Cretaceous)
|
| Nkalagu Borehaole, Eastern Dahomey Basin, Nigeria |
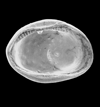
| 1110 microns (length) SEM |
|
| Schuleridea kellawaysii
|
| Bathonian-Callovian (Jurassic)
|
| North Leigh Borehole, Oxfordshire, U.K |
| 590 microns (length) internal view left valve SEM |

|
| Nophrecythere cruciata intermedia
(Lutze)
|
| Callovian (Jurassic)
|
| Cumnor, Oxfordshire, U.K |
| 690 microns (length) left valve SEM |

|
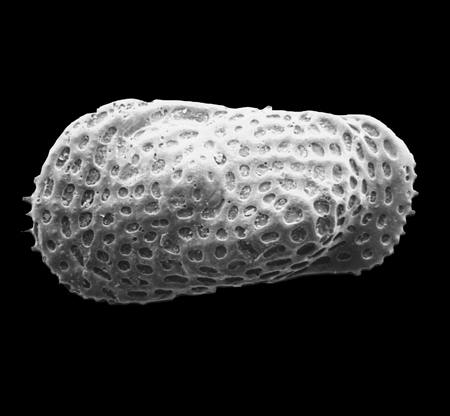
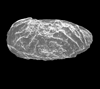
| Polycope plumhoffi
Bate and Coleman
|
| Hettangian-Bathonian (Jurassic)
|
| Trunch Borehole, Norfolk, U.K |
| 250 microns (length) SEM |
|
| Nanacythere minor
Michelsen
|
| Sinemurian (Jurassic)
|
| De Lutte borehole, Eastern Netherlands |
| 310 microns (length) left valve SEM |

|
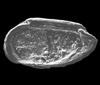
| Gramannicythere bachi bachi
(Gramann)
|
| Sinemurian-Lower Pliensbachian (Jurassic)
|
| Trunch Borehole, Norfolk, U.K |
| 340 microns (length) right valve SEM |
|
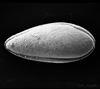
| Gramannicythere bachi bachi
(Gramann)
|
| Sinemurian-Lower Pliensbachian (Jurassic)
|
| Trunch Borehole, Norfolk, U.K |
| 340 microns (length) internal view right valve SEM |
|
| Liasina lanceolata
(Apostolescu)
|
| Upper Sinemurian-Lower Toarcian (Jurassic)
|
| De Lutte borehole, Eastern Netherlands |
| 680 microns (length) right side SEM |
|
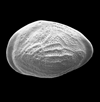
| Kinkelinella intermedia
(Gramann)
|
| Sinemurian (Jurassic)
|
| Trunch Borehole, Norfolk, U.K |
| 640 microns (length) left valve SEM |
|
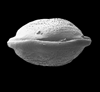
| Kinkelinella intermedia
(Gramann)
|
| Sinemurian (Jurassic)
|
| De Lutte borehole, Eastern Netherlands |
| 450 microns (length) female carapace SEM |
|
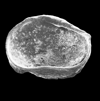
| Rhombocythere penarthensis
Anderson
|
| Norian Rhaetian (Triassic)
|
| Well 6608/11-1 Norweigen North Sea |
| 740 microns (length) internal view right valve SEM |

|
| Rhombocythere penarthensis
Anderson
|
| Norian Rhaetian (Triassic)
|
| Well 6608/11-1 Norweigen North Sea |
| 740 microns (length) left valve SEM |
|