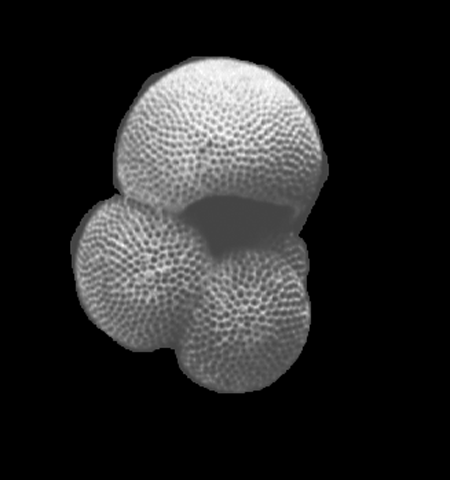
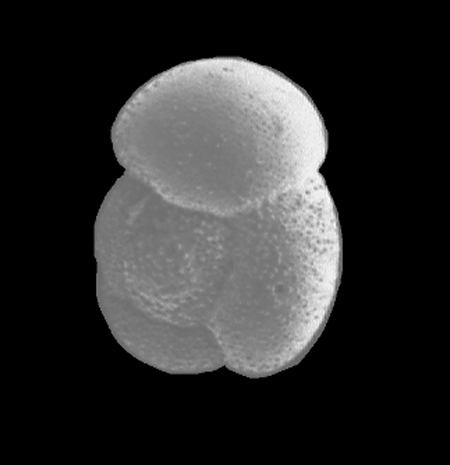
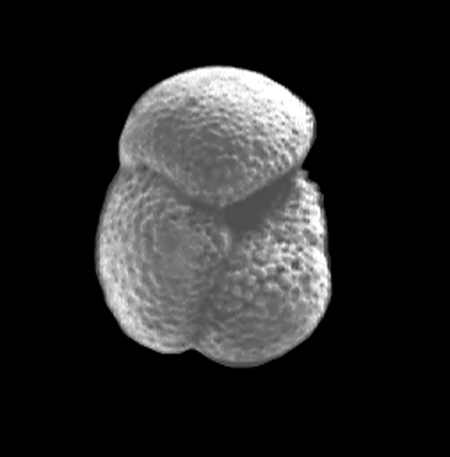
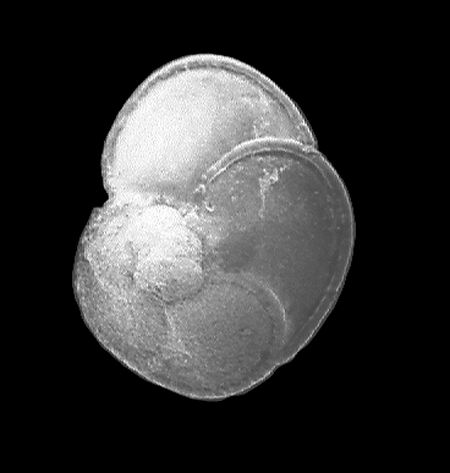
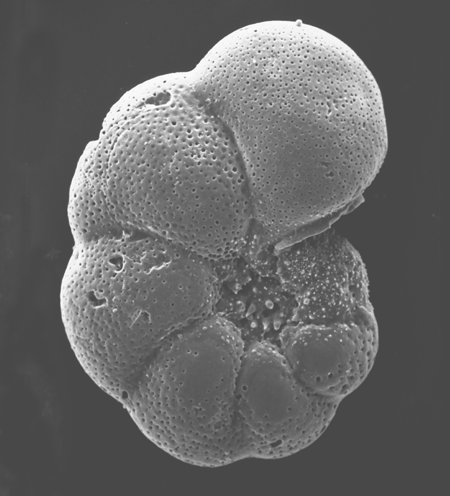
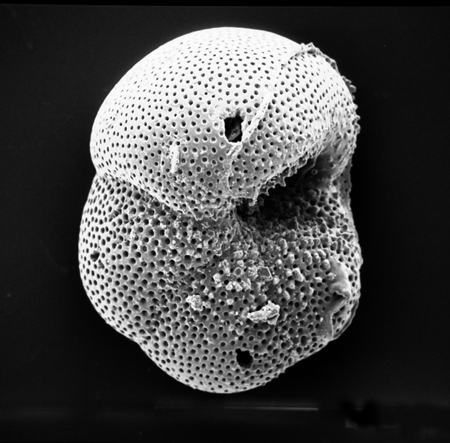
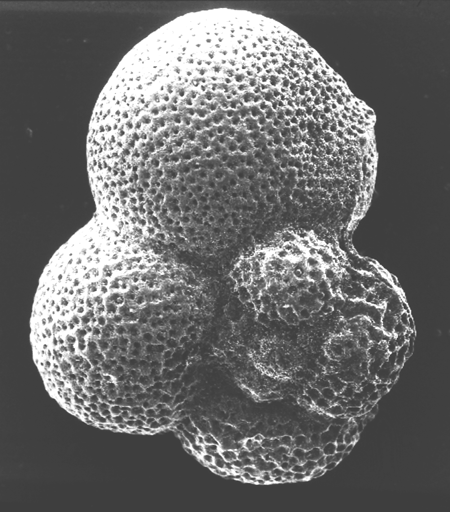
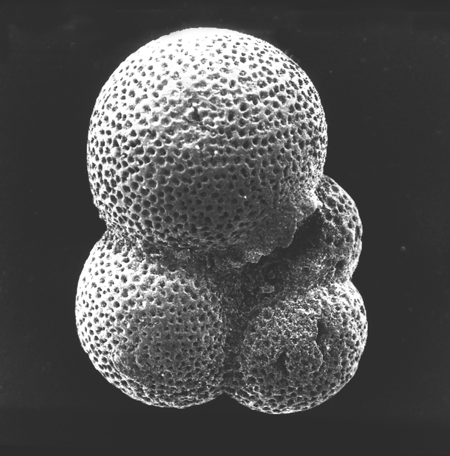
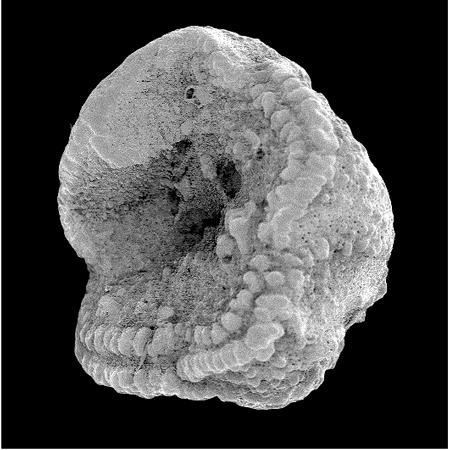
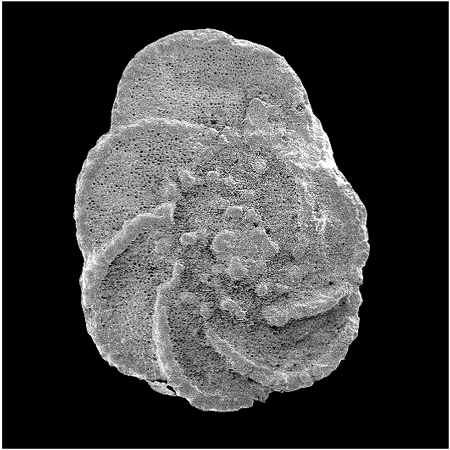
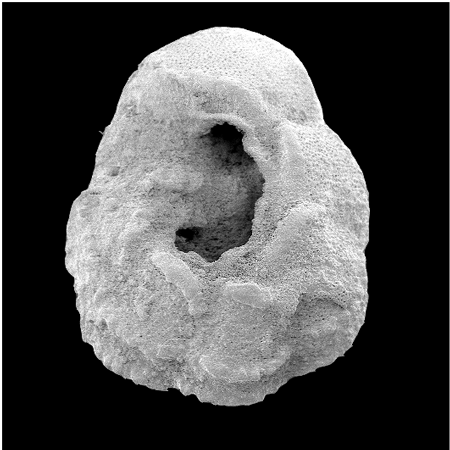
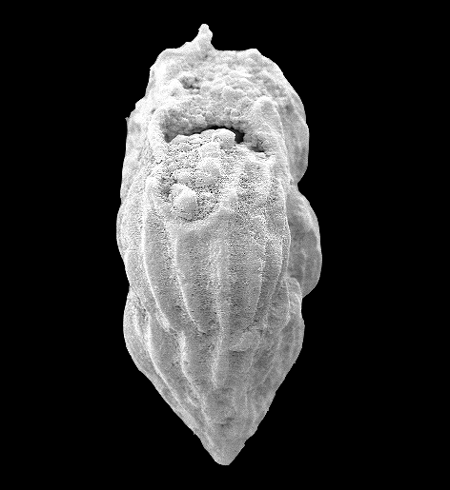
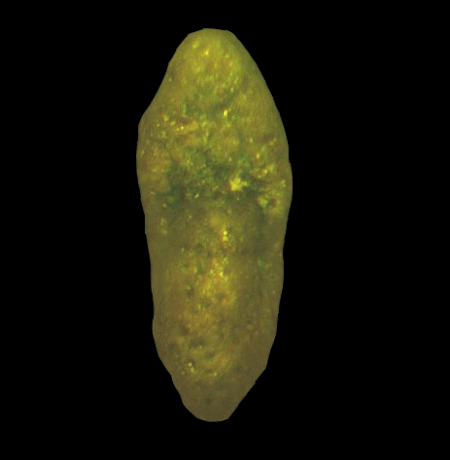
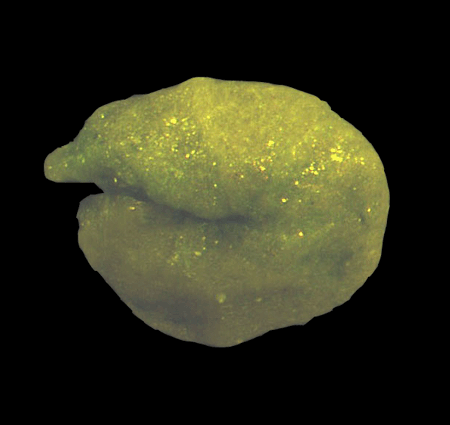
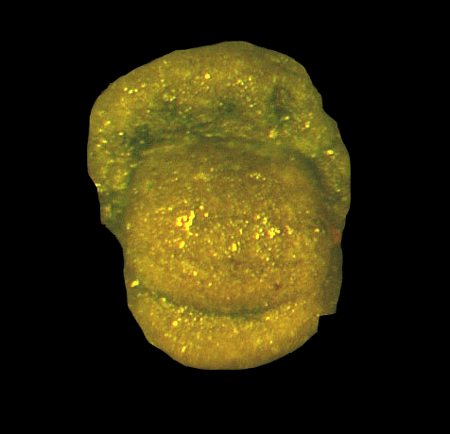
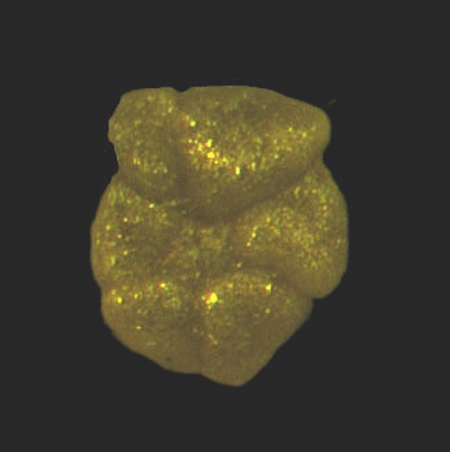
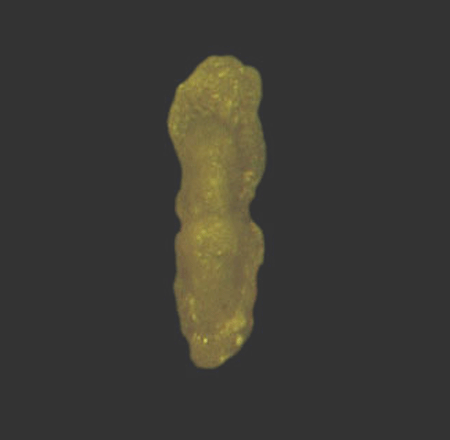
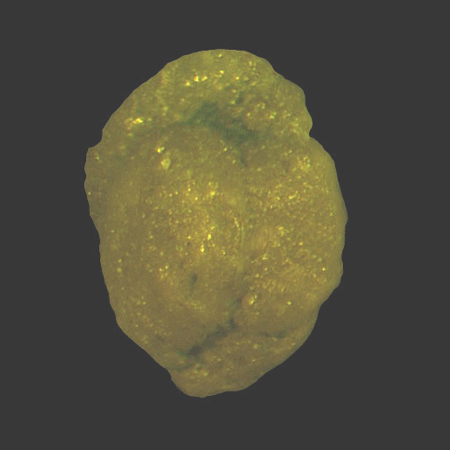
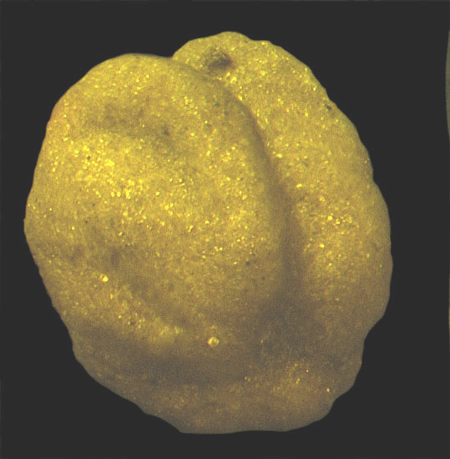
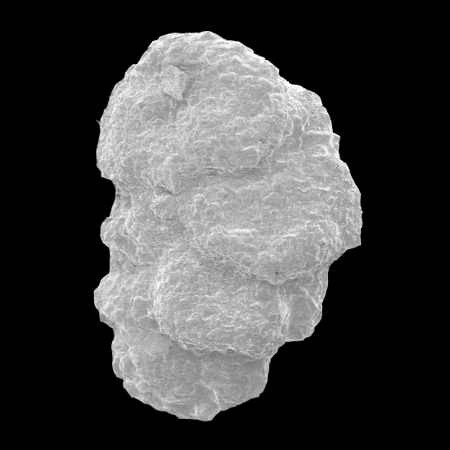
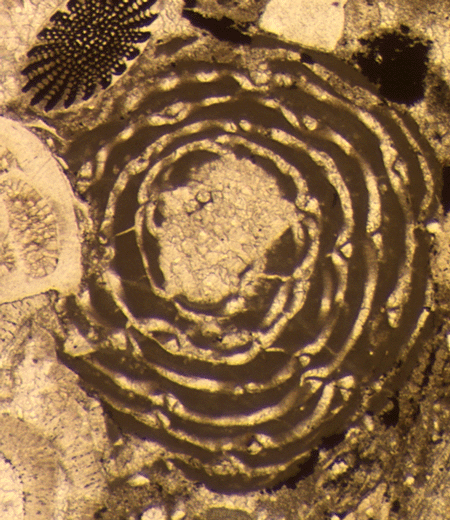
| Introduction |
| Foraminifera are found in all marine environments, they may be planktic or benthic in mode of life. The generally accepted classification of the foraminifera is based on that of Loeblich and Tappan (1964). The Order Foraminiferida (informally foraminifera) belongs to the Kingdom Protista, Subkingdom Protozoa, Phylum Sarcomastigophora, Subphylum Sarcodina, Superclass Rhizopoda, Class Granuloreticulosea. Unpicking this nomenclature tells us that foraminifera are testate (that is possessing a shell), protozoa, (single celled organisms characterised by the absence of tissues and organs), which possess granuloreticulose pseudopodia (these are thread-like extensions of the ectoplasm often including grains or tiny particles of various materials). Bi-directional cytoplasmic flow along these pseudopodia carries granules which may consist of symbiotic dinoflagellates, digestive vacuoles, mitochondria and vacuoles containing waste products; these processes are still not fully understood. In the planktic foraminifera
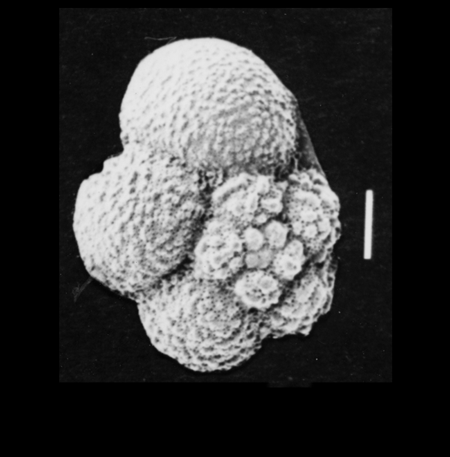
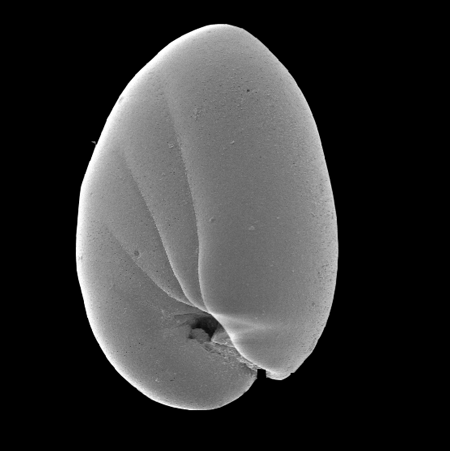
Globigerinoides sacculifer
dinoflagellate symbionts are transported out to the distal parts of rhizopodia in the morning and are returned back into the test at night. The name Foraminiferida is derived from the foramen, the connecting hole through the wall (septa) between each chamber. |
 |
| History of Study |
| The study of foraminifera has a long history, their first recorded "mention" is in Herodotus (fifth century BC) who noted that the limestone of the Egyptian pyramids contained the large benthic foraminifer Nummulites.
In 1835 Dujardin recognised foraminifera as protozoa and shortly afterwards d'Orbigny produced the first classification. The famous
1872 HMS Challenger cruise
, the first scientific oceanographic research expedition to sample the ocean floor collected so many samples that several scientists, including foraminiferologists such as H.B. Brady were still working on the material well in to the 1880's. Work on foraminifera continued throughout the 20th century, workers such as Cushman in the U.S.A and Subbotina in the Soviet Union developed the use of foraminifera as biostratigraphic tools. Later in the 20th century Loeblich and Tappan and Bolli carried out much pioneering work. |
|
| Range |
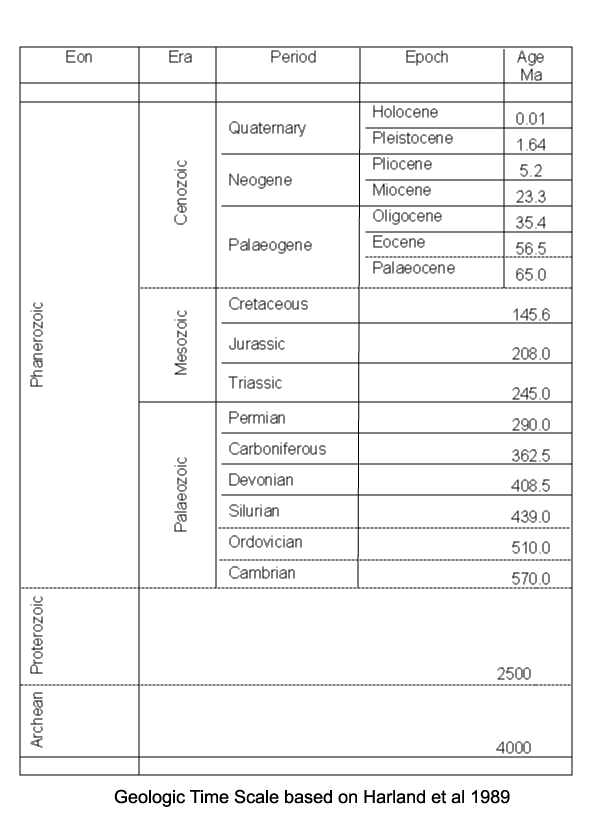
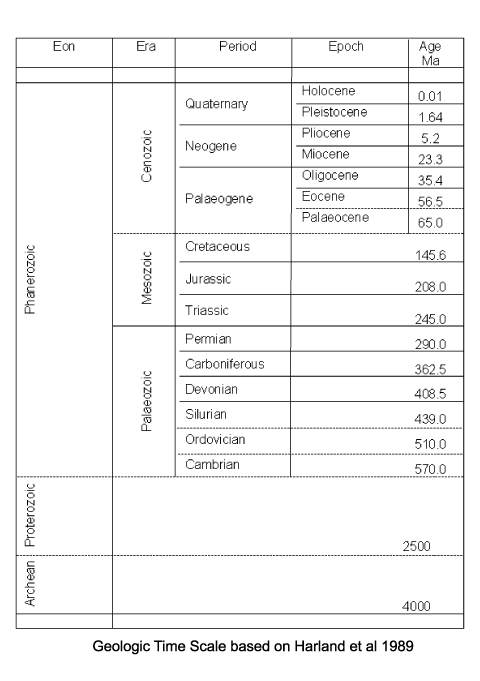
| Foraminifera have a geological range from the earliest Cambrian to the present day. The earliest forms which appear in the fossil record (the allogromiine) have organic test walls or are simple agglutinated tubes. The term "agglutinated" refers to the tests formed from foreign particles "glued" together with a variety of cements. Foraminifera with hard tests are scarce until the Devonian, during which period the fusulinids began to flourish culminating in the complex fusulinid tests of the late Carboniferous and Permian times; the fusulinids died out at the end of the Palaeozoic. The miliolids first appeared in the early Carboniferous, followed in the Mesozoic by the appearance and radiation of the rotalinids and in the Jurassic the textularinids. The earliest forms are all benthic, planktic forms do not appear in the fossil record until the Mid Jurassic in the strata of the northern margin of Tethys and epicontinental basins of Europe. They were probably meroplanktic (planktic only during late stages of their life cycle). The high sea levels and "greenhouse" conditions of the Cretaceous saw a diversification of the planktic foraminifera, and the major extinctions at the end of the Cretaceous included many planktic foraminifera forms. A rapid evolutionary burst occurred during the Palaeocene with the appearance of the planktic globigerinids and globorotalids and also in the Eocene with the large benthic foraminifera of the nummulites, soritids and orbitoids. The orbitoids died out in the Miocene, since which time the large foraminifera have dwindled. Diversity of planktic forms has also generally declined since the end of the Cretaceous with brief increases during the warm climatic periods of the Eocene and Miocene. |
 |
|
| Classification |
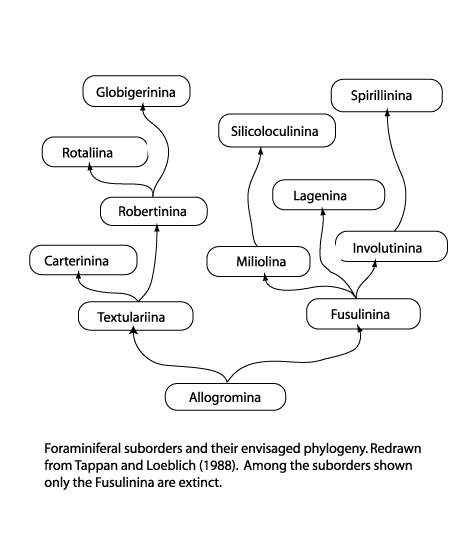
| Foraminifera are classified primarily on the composition and morphology of the test. Three basic wall compositions are recognised, organic (protinaceous mucopolysaccharide i.e. the allogromina), agglutinated and secreted calcium carbonate (or more rarely silica). Agglutinated forms, i.e the Textulariina, may be composed of randomly accumulated grains or grains selected on the basis of specific gravity, shape or size; some forms arrange particular grains in specific parts of the test. Secreted test foraminifera are again subdivided into three major groups, microgranular (i.e. Fusulinina), porcelaneous (i.e. Miliolina) and hyaline (i.e. Globigerinina). Microgranular walled forms (commonly found in the late Palaeozoic) are composed of equidimensional subspherical grains of crystalline calcite. Porcelaneous forms have a wall composed of thin inner and outer veneers enclosing a thick middle layer of crystal laths, they are imperforate and made from high magnesium calcite. The hyaline foraminifera add a new lamella to the entire test each time a new chamber is formed; various types of lamellar wall structure have been recognised, the wall is penetrated by fine pores and hence termed perforate. A few "oddities" are also worth mentioning, the Suborder Spirillinina has a test constructed of an optically single crystal of calcite, the Suborder Silicoloculinina as the name suggests has a test composed of silica. Another group (the Suborder Involutina) have a two chambered test composed of aragonite. The Robertinina also have a test composed of aragonite and the Suborder Carterina is believed to secrete spicules of calcite which are then weakly cemented together to form the test. |
 |
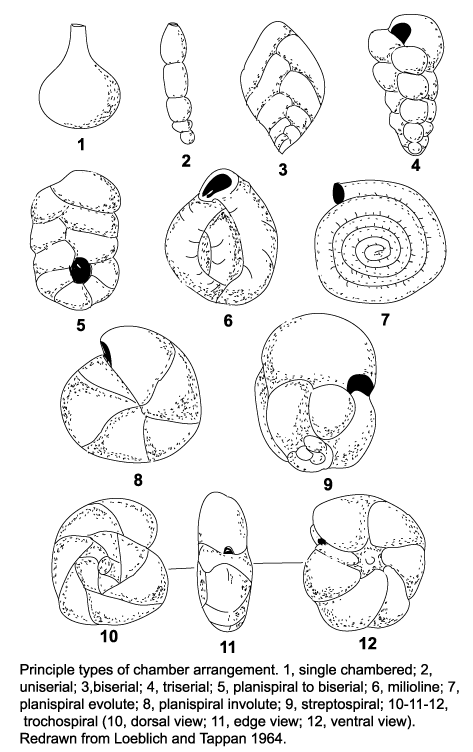
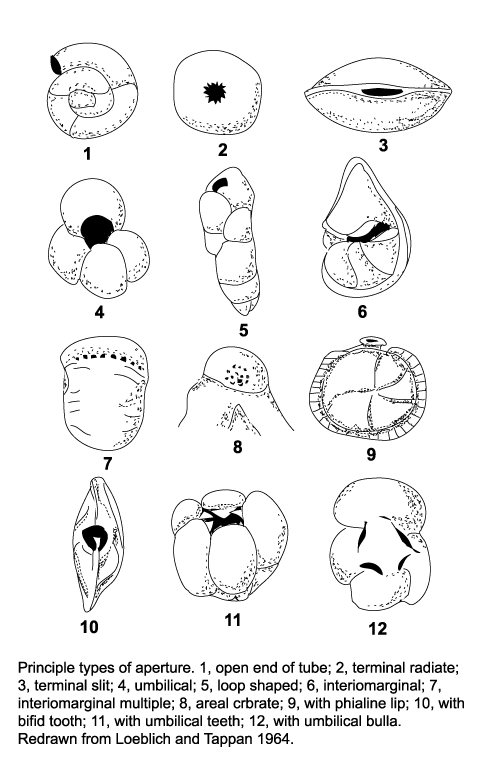
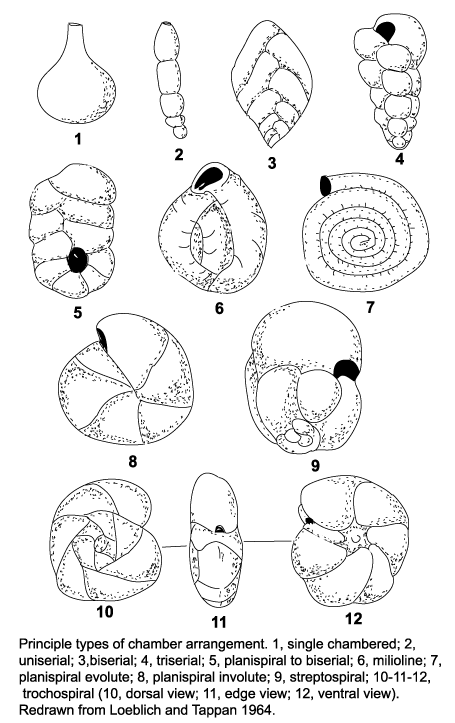
| The morphology of foraminifera tests varies enormously, but in terms of classification two features are important. Chamber arrangement and aperture style, with many subtle variations around a few basic themes. These basic themes are illustrated in the following two diagrams but it should be remembered that these are only the more common forms and many variations are recognised. |
 |
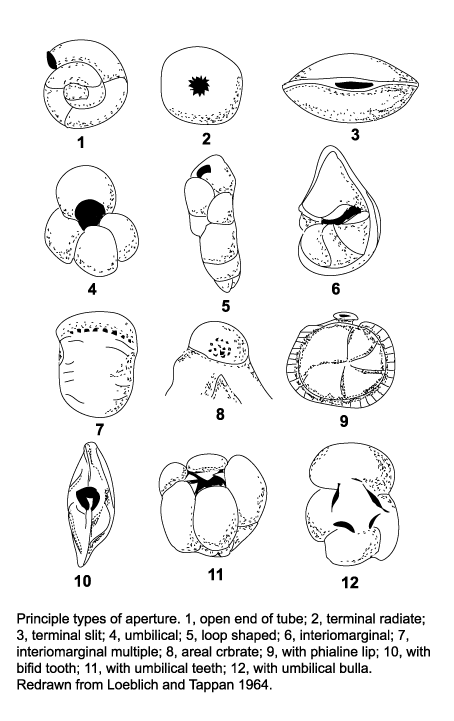 |
|
| Applications |
| As previously mentioned, foraminifera have been utilised for biostratigraphy for many years, and they have also proven invaluable in palaeoenvironmental reconstructions most recently for palaeoceanographical and palaeoclimatological purposes. For example palaeobathymetry, where assemblage composition is used and palaeotemperature where isotope analysis of foraminifera tests is a standard procedure. In terms of biostratigraphy, foraminifera have become extremely useful, different forms have shown evolutionary bursts at different periods and generally if one form is not available to be utilised for biostratigraphy another is. For example preservation of calcareous walled foraminifera is dependent on the depth of the water column and Carbonate Compensation Depth (the depth below which dissolution of calcium carbonate exceeds the rate of its deposition), if calcareous walled foraminifera are therefore not preserved agglutinated forms may be. The oldest rocks for which foraminifera have been biostratigraphically useful are Upper Carboniferous to Permian strata, which have been zoned using the larger benthic fusulinids. Planktic foraminifera have become increasingly important biostratigraphic tools, especially as petroleum exploration has extended to offshore environments of increasing depths. The first and last occurrence of distinctive "marker species" from the Cretaceous to Recent (particularly during the Upper Cretaceous) has allowed the development of a well established fine scale biozonation. |
| Benthic foraminifera have been used for palaeobathymetry since the 1930's and modern studies utilise a variety of techniques to reconstruct palaeodepths. For studies of relatively recent deposits simple comparison to the known depth distribution of modern extant species is used. For older material changes in species diversity, planktic to benthic ratios, shell-type ratios and test morpholgy have all been utilised. Variations in the water temperature inferred from oxygen isotopes from the test calcite can be used to reconstruct palaeoceanographic conditions by careful comparison of changes in oxygen isotope levels as seen in benthic forms (for bottom waters) and planktic forms(for mid to upper waters). This type of study has allowed the reconstruction of oceanic conditions during the Eocene-Oligocene, the Miocene and the Quaternary. Benthic foraminifera have been divided into morphogroups based on the test shape and these groups used to infer palaeo-habitats and substrates; infaunal species tending to be elongate and streamlined in order to burrow into the substrate and epifaunal species tending to be more globular with one relatively flatter side in order to facilitate movement on top of the substrate. It should be remembered, however, that a large variety of morphologies and possible habitats have been recognised making such generalisations of only limited use. Studies of modern foraminifera have recognised correlations between test wall type (for instance porcelaneous, hyaline, agglutinated), palaeodepths and salinity by plotting them onto triangular diagrams. |
|
| Biology |
| Studies of living foraminifera, in controlled laboratory environments, have provided limited information regarding trophic strategies but much has been inferred by relating test morphology to habitat. Foraminifera utilise a huge variety of feeding mechanisms, as evidenced by the great variety of test morphologies that they exhibit. From the variety of trophic habits and test morphologies a few generalisations may be made. Branching benthic foraminifera such as
Notodendrodes antarctikos
,which resembles a microscopic tree, absorbs dissolved organic matter via a "root" system. Other sessile benthic foraminifera exhibit test morphologies dependent on the substrate on or in which they live, many are omnivorous opportunistic feeders and have been observed to consume autotrophic and heterotrophic protists (including other foraminifera), metazoans and detritus. Some suspension feeding foraminifera utilise their pseudopodia to capture food from the water column, or interstitial pore waters,
Elphidium crispum
forms a "spiders web" between the stipes of coralline algae. Infaunal forms are probably detritivores and commonly have elongate tests to facilitate movement through the substrate. Benthic and planktonic foraminifera which inhabit the photic zone often live symbiotically with photosynthesising algae such as dinoflagellates, diiatoms and chlorophytes. It is thought the large benthic, discoidal and fusiform foraminifera attain their large size in part because of such associations. Foraminifera are preyed upon by many different organisms including worms, crustacea, gastropods, echinoderms, and fish. It should be remembered that the biocoenosis (life assemblage) will be distorted by selective destruction by predators. |
|
| Life Cycle |
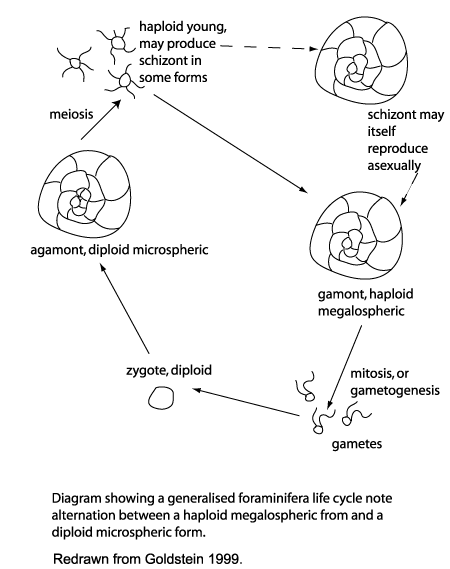 |
| Of the approximately 4000 living species of foraminifera the life cycles of only 20 or so are known. There are a great variety of reproductive, growth and feeding strategies, however the alternation of sexual and asexual generations is common throughout the group and this feature differentiates the foraminifera from other members of the Granuloreticulosea. An asexually produced haploid generation commonly form a large proloculus (initial chamber) and are therefore termed megalospheric. Sexually produced diploid generations tend to produce a smaller proloculus and are therefore termed microspheric. Importantly in terms of the fossil record, many foraminiferal tests are either partially dissolved or partially disintegrate during the reproductive process.The planktonic foraminifera
Hastigerina pelagica
reproduces by gametogenesis at depth, the spines, septa and apertural region are resorbed leaving a tell-tale test.
Globigerinoides sacculifer
produces a sac-like final chamber and additional calcification of later chambers before dissolution of spines occurs, this again produces a distinctive test, which once gametogenesis is complete sinks to the sea bed. |
|
| Preparation Techniques |
|
WARNING:
Please remember all preparation techniques require the use of hazardous materials and equipment and should only be carried out in properly equiped laboratories, wearing the correct safety clothing and under the supervision of qualified staff. |
| Foraminifera range in size from several millimeters to a few tens of microns and are preserved in a variety of rock types. The preparation techniques used depend on the rock type and the "predicted" type of foraminifera one expects to find. Very hard rocks such as many limestones are best thin sectioned as in normal petrological studies, except instead of grinding to a set thickness (commonly 30 microns) the sample is ground very carefully by hand until the optimum thickness is obtained for each individual sample. This is a skilled job and requires expensive equipment but provides excellent results and is particularly used in the study of larger benthic foraminifera from reef type settings. |
| Planktic and smaller benthic foraminifera are prepared by crushing the sample into roughly five millimeter fragments. The crushed sample is then placed in a strong glass beaker or similar vessel and water and washing soda or 6% hydrogen peroxide added, left to stand and then heated and allowed to simmer. The length of time the sample is left to simmer depends on the rock type involved and if peroxide is used the sample should not be left immersed in the solution for more than about half an hour. Next, the material is washed through a 63 micron sieve untill the liquid coming through the sieve is clean (i.e. the clay fraction has been removed). The sample can then be dried and sieved into fractions (generally 63-125 microns, 125-250 microns, 250-500 microns and greater than 500 microns) using a "nest" of dry sieves. Care must be taken to clean all sieves and materials used between the preparation of each sample to prevent contamination. |
|
| Observation Techniques |
| Thin sections are veiwed using transmitted-light petrological type microscopes. Washed, dried fossil samples can be picked from any remaining sediment using a fine brush and a reflected light, binocular microscope. The best method is to scatter a fine dusting of sieved sediment on to a black tray divided into squares, this can then be scanned under the microscope and any foraminifera preserved in the sediment can be picked out with a fine brush (preferably a 000 sable-haired brush). The picked specimens can then be mounted in card slides divided into numbered squares with sliding glass covers. Gum tragocanth was traditionally used to attach the specimens to the slides but modern office-type paper adhesives are now used. |
|
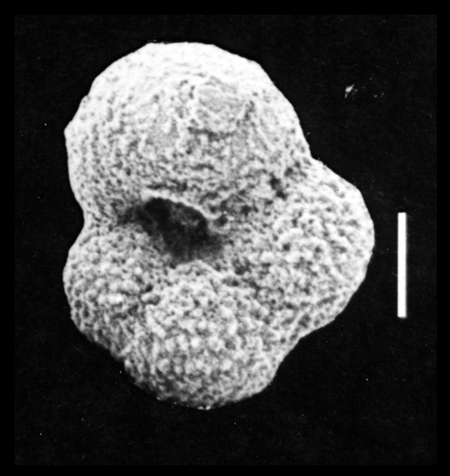
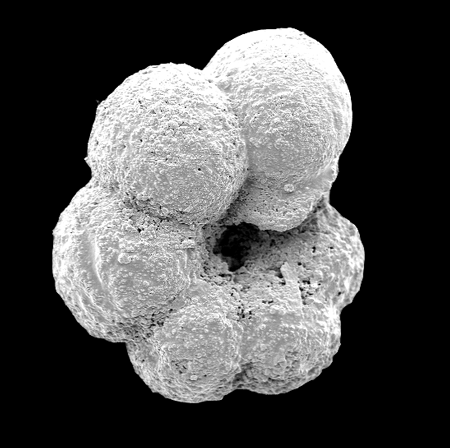
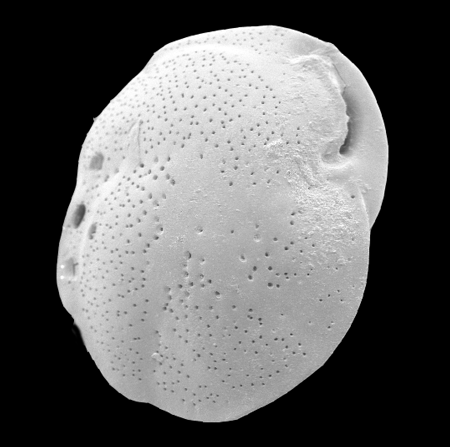
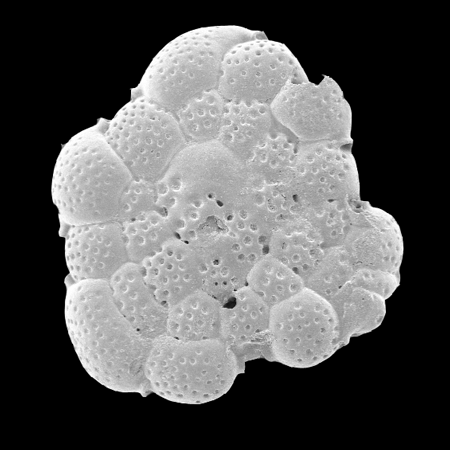
| Images |
| The following images are of a representative selection of foraminifera aimed at giving a general overview of the different morphotypes. Each specimen is given a generic and, if possible, a species name followed by its age range, the site location from which the sample was obtained and its size in microns. LM (Light Microscope) SEM (Scanning Electron Microscope) TS (Thin Section). Typical and selected marker species are illustrated from each main period of the geological column in which foraminifera occur. Click on an image to view a larger version. Because Foraminifera formsuch a diverse taxon they have been split into three groups: planktics, benthics and larger benthics. |
|
|
| Planktic |

|
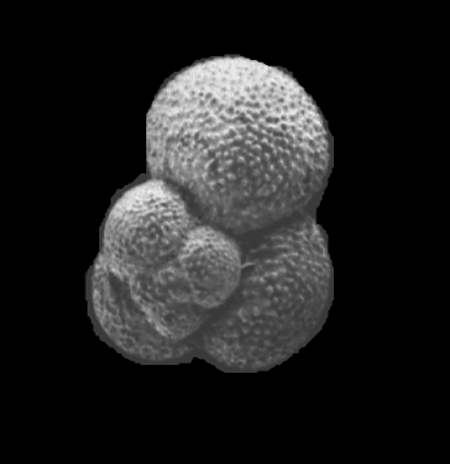
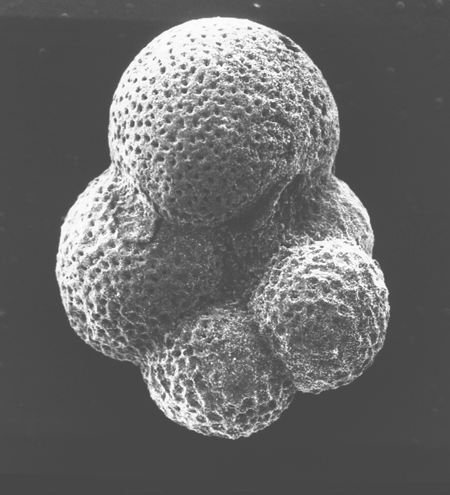
| Globigerina bulloides
d'Orbigny
|
| Pliocene-Recent
|
| South Africa |
| 380 microns spiral view SEM |

|
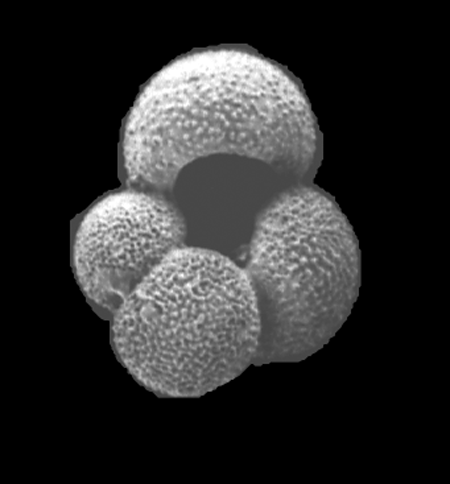
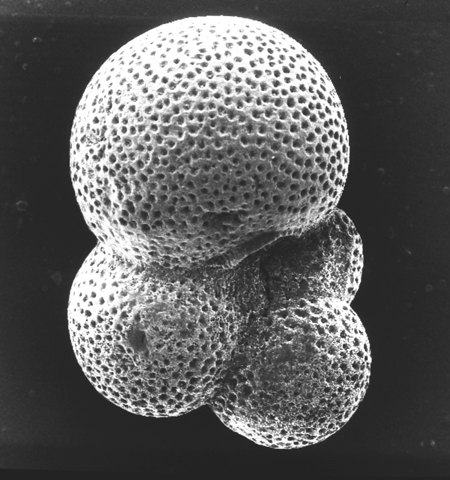
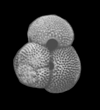
| Globigerina bulloides
d'Orbigny
|
| Pliocene-Recent
|
| South Africa |
| 416 microns umbilical view SEM |
|
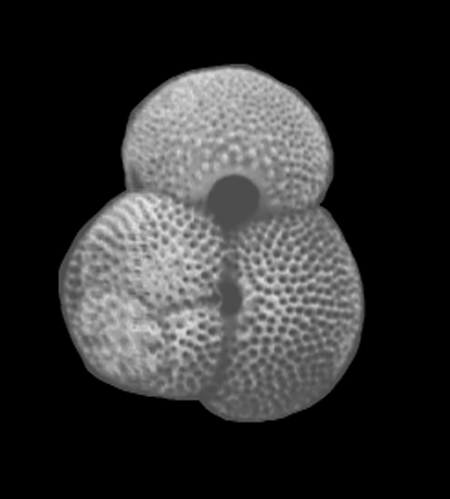
| Globigerinoides ruber
d'Orbigny
|
| Miocene-Recent
|
| South Africa |
| spiral view SEM |

|
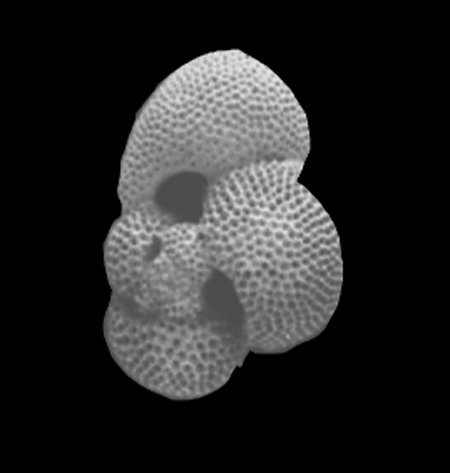
| Globigerinoides sacculifer
(Brady)
|
| Miocene-Recent
|
| South Africa |
| spiral view SEM |

|
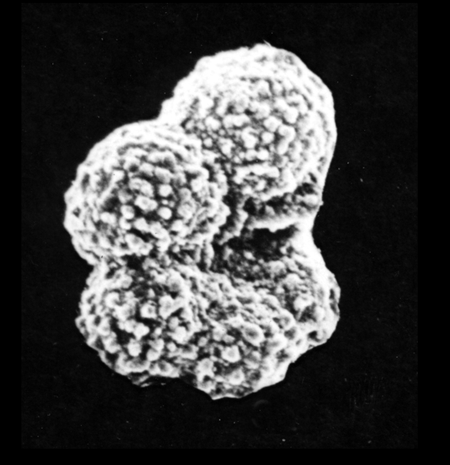
| Globigerinoides sacculifer
(Brady)
|
| Miocene-Recent
|
| South Africa |
| umbilical view SEM |

|
| Globorotalia inflata
d'Orbigny
|
| Pliocene-Recent
|
| South Africa |
| spiral view SEM |

|
| Globorotalia inflata
d'Orbigny
|
| Pliocene-Recent
|
| South Africa |
| umbilical view SEM |

|
| Globorotalia menardii
(Parker, Jones and Brady)
|
| Pliocene-Recent
|
| South Africa |
| spiral view SEM |

|
| Globorotalia menardii
(Parker, Jones and Brady)
|
| Pliocene-Recent
|
| South Africa |
| umbilical view SEM |

|
| Neogloboquadrina pachyderma
(Ehrenberg)
|
| Pliocene-Recent
|
| South Africa |
| spiral view SEM |

|
| Neogloboquadrina pachyderma
(Ehrenberg)
|
| Pliocene-Recent
|
| South Africa |
| umbilical view SEM |

|
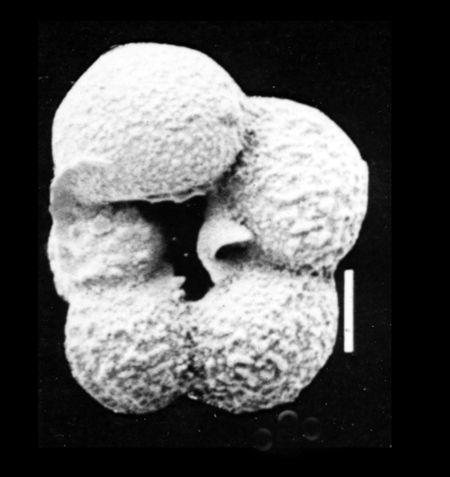
| Orbulina universa
d'Orbigny
|
| Middle Miocene-Recent
|
| South Africa |
| SEM |

|
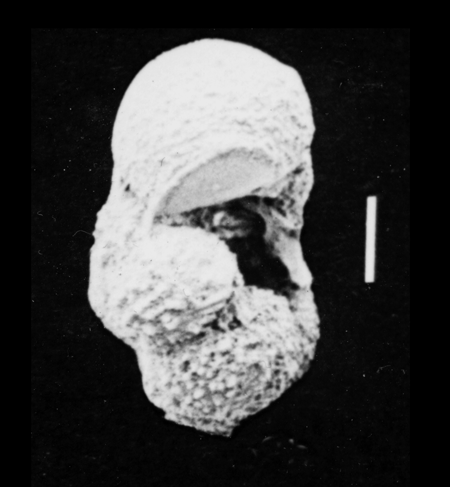
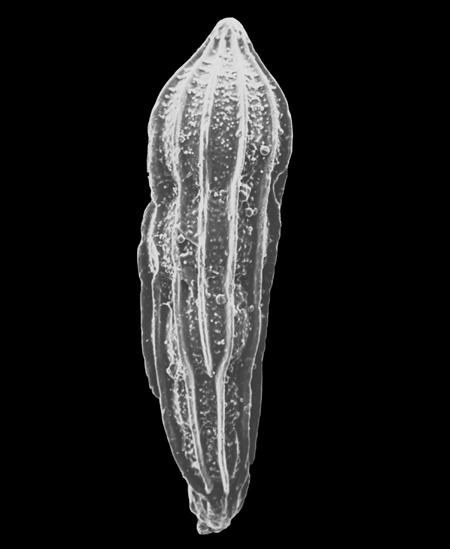
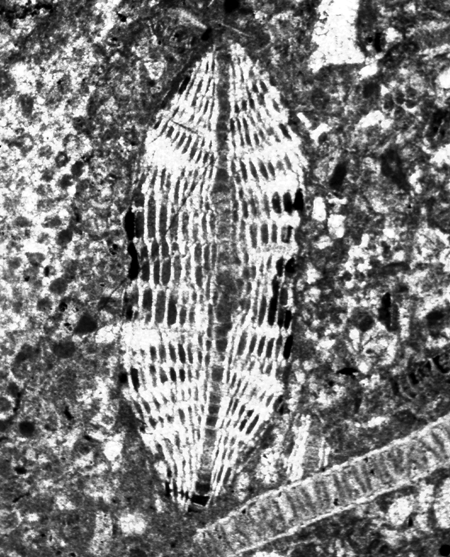
| Hantkenina alabamensis
Cushman, 1927
|
| Eocene
|
| Montgomery Landing, Red River, Louisiana, USA |
| side view (slightly broken specimen) SEM |

|
| Pseudohastigerina micra
(Cole, 1927)
|
| Eocene-Oligocene
|
| Montgomery Landing, Red River, Louisiana, USA |
| side view SEM |

|
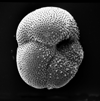
| Globorotalia centralis
Cushman and Bermudez, 1937
|
| Eocene
|
| Montgomery Landing, Red River, Louisiana, USA |
| umbilical view SEM |
|
| Globorotalia cerro-azulensis
Cole, 1928
|
| Eocene
|
| Montgomery Landing, Red River, Louisiana, USA |
| umbilical view SEM |

|
| Parasubbotina varianta
(Subbotina, 1953)
|
| Lower-Middle Palaeocene
|
| Zin Valley, Israel |
| spiral view SEM |

|
| Parasubbotina varianta
(Subbotina, 1953)
|
| Lower-Middle Palaeocene
|
| Zin Valley, Israel |
| umbilical view SEM |

|
| Parasubbotina pseudobulloides
(Plummer, 1926)
|
| Lower-Middle Palaeocene
|
| Zin Valley, Israel |
| umbilical view SEM |

|
| Subbotina triloculinoides
(Plummer, 1926)
|
| lower-upper Palaeocene
|
| Zin Valley, Israel |
| umbilical view SEM |

|
| Subbotina triloculinoides
(Plummer, 1926)
|
| Palaeocene
|
| Zin Valley, Israel |
| spiral view SEM |

|
| Abathomphalus mayaroensis
(Bolli)
|
| Upper Maastrichtian (Upper Cretaceous)
|
| Kassbah, N.W. Syria |
| umbilical view SEM |

|
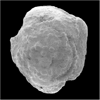
| Abathomphalus mayaroensis
(Bolli)
|
| Upper Maastrichtian (Upper Cretaceous)
|
| Kassbah, N.W. Syria |
| lateral view SEM |
|
| Contusotruncana contusa
(Cushman)
|
| Upper Maastrichtian (Upper Cretaceous)
|
| Kassbah, N.W. Syria |
| dorsal view SEM |

|
| Contusotruncana contusa
(Cushman)
|
| Upper Maastrichtian (Upper Cretaceous)
|
| Kassbah, N.W. Syria |
| ventral view SEM |

|
| Globotruncana linneiana
(d'Orbigny)
|
| Santonian-Maastrichtian (Upper Cretaceous)
|
| Kassbah, N.W. Syria |
| dorsal view SEM |

|
| Globotruncana linneiana
(d'Orbigny)
|
| Santonian-Maastrichtian (Upper Cretaceous)
|
| Kassbah, N.W. Syria |
| ventral view SEM |

|
| Racemiguembelina fructicosa
(Egger)
|
| Middle-Uppper Maastrichtian (Upper Cretaceous)
|
| Kassbah, N.W. Syria |
| SEM |

|
| Racemiguembelina fructicosa
(Egger)
|
| Middle-Uppper Maastrichtian (Upper Cretaceous)
|
| Kassbah, N.W. Syria |
| SEM |

|
| Pseudotextularia elegans
(Rzehak)
|
| Campanian-Maastrichtian (Upper Cretaceous)
|
| Kassbah, N.W. Syria |
| SEM |

|
| Pseudoguembelina excolata
(Cushman)
|
| Campanian-Maastrichtian (Upper Cretaceous)
|
| Kassbah, N.W. Syria |
| SEM |

|
| Archaeoglobigerina cretacea
(d'Orbigny)
|
| Coniacian-Maastrichtian (Upper Cretaceous)
|
| Sens, N. France |
| scale bar 100 microns edge view SEM |

|
| Archaeoglobigerina cretacea
(d'Orbigny)
|
| Coniacian-Maastrichtian (Upper Cretaceous)
|
| Sens, N. France |
| scale bar 100 microns dorsal view SEM |

|
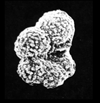
| Archaeoglobigerina cretacea
(d'Orbigny)
|
| Coniacian-Maastrichtian (Upper Cretaceous)
|
| Sens, N. France |
| scale bar 100 microns ventral view SEM |
|
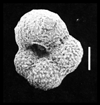
| Hedbergella delrioensis
(Carsey)
|
| Coniacian-Santonian (Upper Cretaceous)
|
| Faircross, UK |
| scale bar 100 microns ventral view SEM |
|
| Whiteinella baltica
Douglas and Rankin
|
| Coniacian-Santonian (Upper Cretaceous)
|
| Winterbourne, UK |
| scale bar 100 microns ventral view SEM |

|
| Heterohelix pulchra
(Brotzen)
|
| Coniacian-Maastrichtian (Upper Cretaceous)
|
| N. Norfolk, UK |
| scale bar 100 microns side view SEM |

|
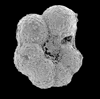
| Heterohelix globulosa
(Ehrenberg)
|
| Coniacian-Maastrichtian (Upper Cretaceous)
|
| Sens, N. France |
| scale bar 100 microns side view SEM |
|
| Hedbergella planispira
(Tappan)
|
| Aptian-Coniacian (Upper Cretaceous)
|
| Karai, S.E. India |
| ventral view SEM |

|
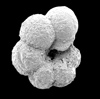
| Hedbergella sigali
Moullade
|
| Barremian-Aptian (Lower Cretaceous)
|
| Karai, S.E. India |
| ventral view SEM |
|
| Ticinella primula
Luterbacher
|
| Albian (Lower Cretaceous)
|
| Karai, S.E. India |
| ventral view SEM |
|
| Benthic |

|
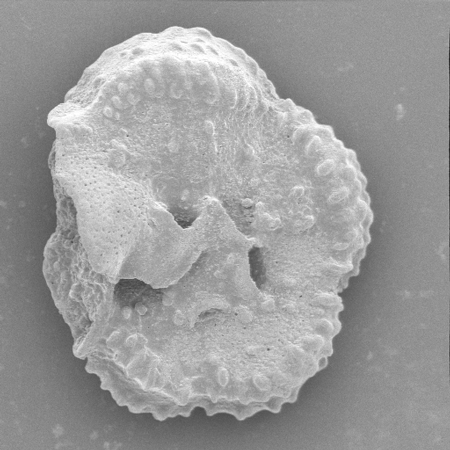
| Spiroloculina ornata
(d'Orbigny)
|
| -Recent
|
| Sea of Marmara |
| side view SEM |

|
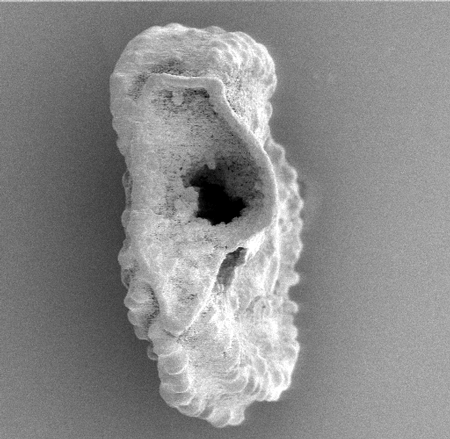
| Elphidium macellum
(Fichtel and Moll)
|
| -Recent
|
| Sea of Marmara |
| side view SEM |

|
| Brizalina alata
(Seguenza)
|
| -Recent
|
| Sea of Marmara |
| side view SEM |

|
| Cassidulina neocarinata
(Thalmann)
|
| -Recent
|
| Sea of Marmara |
| ventral view SEM |

|
| Siphotextularia concava
(Karrer)
|
| -Recent
|
| Sea of Marmara |
| side view SEM |

|
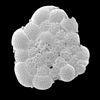
| Bigenerina nodosaria
(d'Orbigny)
|
| ??-Recent
|
| Sea of Marmara |
| side view SEM |
|
| Planorbulina mediterranensis
(d'Orbigny)
|
| ??-Recent
|
| Sea of Marmara |
| unattached side SEM |

|
| Nonionella opima
(Cushman)
|
| ??-Recent
|
| Sea of Marmara |
| side view SEM |

|
| Lagena striata
(d'Orbigny)
|
| ??-Recent
|
| Sea of Marmara |
| side view SEM |

|
| Alveovalvulina suteri
Bronnimann
|
|
|
| Gulf of Mexico |
| LM |

|
| Cyclamina cancellata
Brady
|
|
|
| Gulf of Mexico |
| 1000 microns LM |

|
| Cyclamina acutidorsata
(von Hantken)
|
| Oligocene-Miocene
|
| Gulf of Mexico |
| 1000 microns LM |

|
| Cyclamina acutidorsata
(von Hantken)
|
|
|
| Gulf of Mexico |
| 1000 microns LM |

|
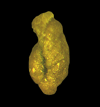
| Reticulophragmium rotundidorsata
(von Hantken)
|
| Eocene-Miocene
|
| Gulf of Mexico |
| 600 microns LM |
|
| Cribrostomoides
sp.
|
|
|
| Gulf of Mexico |
| 600 microns side view LM |

|
| Cribrostomoides
sp.
|
|
|
| Gulf of Mexico |
| 600 microns apertural view LM |

|
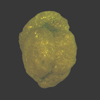
| Haplophragmoides bradyi
(Robertson)
|
|
|
| Gulf of Mexico |
| 600 microns side view LM |

|
| Haplophragmoides bradyi
(Robertson)
|
|
|
| Gulf of Mexico |
| 600 microns apertural view LM |
|
| Jarvisella karamatensis
Bronnimann
|
|
|
| Gulf of Mexico |
| 600 microns apertural view LM |

|
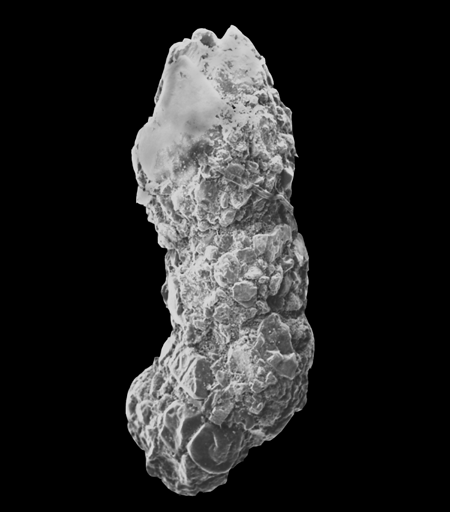
| Recurvoides azuamensis
Bermudez
|
| Oligocene?
|
| Gulf of Mexico |
| 1500 microns apertural view LM |

|
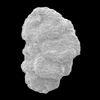
| Textularia tatumi
Cushman and Ellisor
|
| Miocene?
|
| Gulf of Mexico |
| SEM |
|
| Valvulina flexilis
Cushman and Renz
|
| Oligocene-Miocene
|
| Gulf of Mexico |
| SEM |

|
| Nodosaria
sp.
|
| Jurassic
|
| Villers sur Mer, Normandy, France |
| SEM |

|
| Nodosaria
sp.
|
| Jurassic
|
| Villers sur Mer, Normandy, France |
| close up of aperture SEM |

|
| Vaginulina bernardi
Paalzow
|
| Oxfordian (Jurassic)
|
| Villers sur Mer, Normandy, France |
| SEM |

|
| Ammobaculites coprolithiformis
Schwager
|
| Bathonian-Kimmeridgian (Jurassic)
|
| Villers sur Mer, Normandy, France |
| SEM |

|
| Psammosphaera
sp. Schulze
|
| middle Ordovician-Recent
|
| Voring Basin, offshore Norway |
| SEM |

|
| Miliamina fusca
(Bradey)
|
| ?-Recent
|
| Voring Basin, offshore Norway |
| SEM |

|
| Quinqueloculina impressa
Reuss
|
| ?-Eocene
|
| Voring Basin, offshore Norway |
| SEM |
|
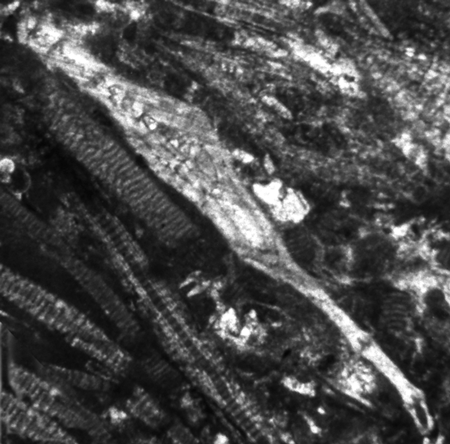
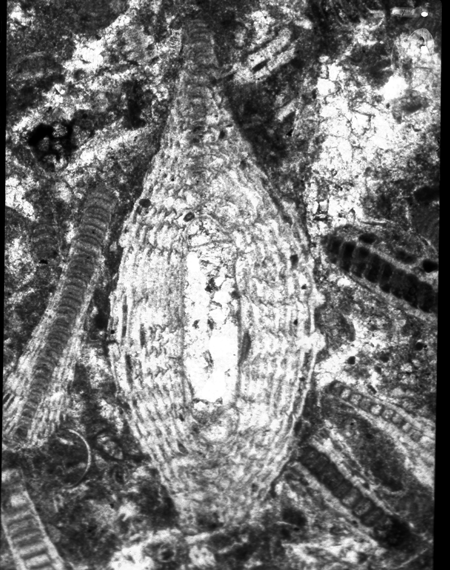
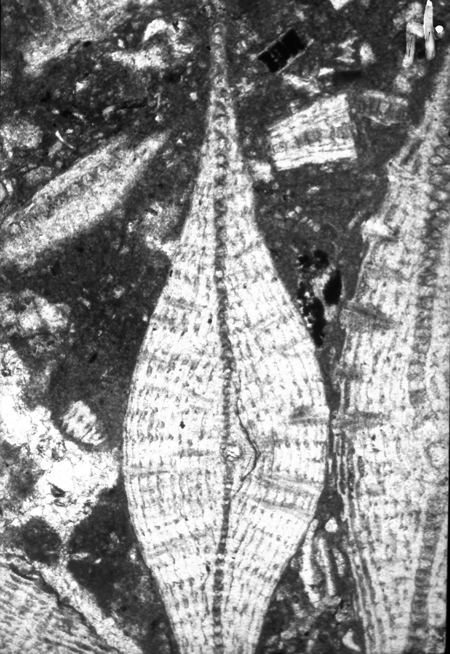
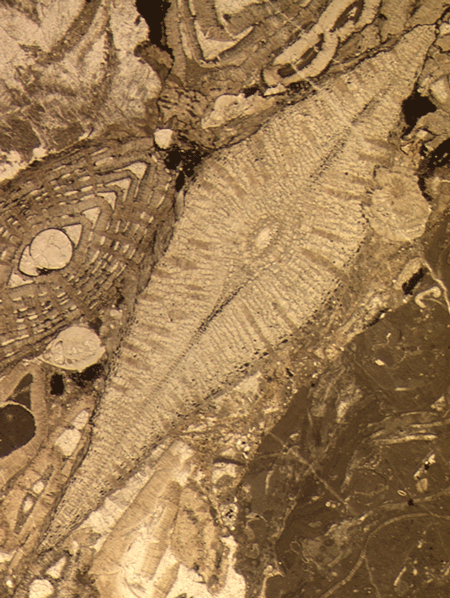
| Larger Benthic |

|
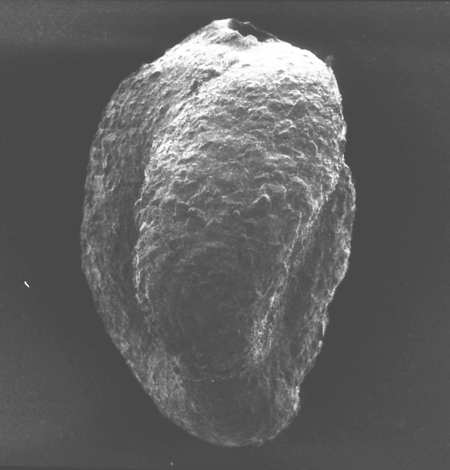
| Helicolepidina cf nortoni
Vaughan
|
| ?Eocene
|
| Barinas, S.W Venezuela |
| Thin Section |

|
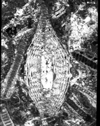
| Heterostegina
sp d'Orbigny
|
| ?Eocene
|
| Barinas, S.W Venezuela |
| TS |
|
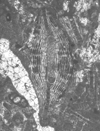
| Lepidocyclina tobleri panamensis
(Cushman)
|
| ?Eocene
|
| Barinas, S.W Venezuela |
| TS |
|
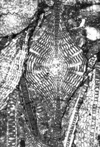
| Lepidocyclina pustulosa tobleri
(Douville)
|
| ?Eocene
|
| Barinas, S.W Venezuela |
| TS |
|
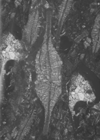
| Lepidocyclina pustulosa pustulosa
(Douville)
|
| ?Eocene
|
| Barinas, S.W Venezuela |
| TS |
|
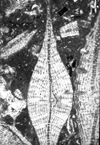
| Discocyclina (Discocyclina) marginata
(Cushman)
|
| ?Eocene
|
| Barinas, S.W Venezuela |
| TS |
|
| Lepidocyclina cf tobleri
(Cushman)
|
| ?Eocene
|
| Barinas, S.W Venezuela |
| TS |

|
| Nummulites
sp. Lamarck
|
| Paleocene-Holocene
|
| Ainsa, Southern Pyrennees, Spain |
| Thin section |

|
| Discocyclina
sp. Gumbel
|
| Mid Paleocene-Upper Eocene
|
| Ainsa, Southern Pyrennees, Spain |
| Thin section |

|
| Alveolina
sp. d'Orbigny
|
| Upper Paleocene-Upper Eocene
|
| Ainsa, Southern Pyrennees, Spain |
| Thin section |

|
| Alveolina
sp. d'Orbigny
|
| Upper Paleocene-Upper Eocene
|
| Ainsa, Southern Pyrennees, Spain |
| Thin section |
|