
Social evolution and kin selectionThe lecture will be illustrated with examples, so if you miss the lecture, you'll miss the pix! For copyright reasons, I can't show all the pictures in the web notes; only some of my own.
Linkage disequilibria and evolution at more than one gene
Evolutionary developmental genetics
The involvement of genetic drift in evolution, including the shifting balance
Race formation and speciation
Biogeography
Warning colour and mimicry form a part of the general topic of defensive coloration, with much of which you will already be familiar (if not fed up to the back teeth of, because of overexposure to books and TV films as a kid). So we can zip through the whole area of defensive coloration quickly.
Individuals avoid visually-hunting predators, or, if they ARE predators, avoid being detected by visually aware prey using the following techniques:
Camouflage - also called crypsis, cryptic coloration. Individuals rest, and have coloration and patterns similar to those in the visual background.Flash coloration. Individuals are cryptic at first. When prodded, they may fly or jump about, suddenly flashing brightly-coloured appendages or underwings normally hidden at rest. This is very common in certain flying grasshoppers which have blue or red underwings. It is hypothesised that predators are distracted and search for blue or red objects, which of course have disappeared when the animal settles again.
In some cases, frightening eyespots or bright red/yellow/blue markings appear, particularly on the underwings of Sphingidae (hawk-moths) and Noctuidae: Catacolinae (underwing moths). Birds encountering these prey are startled and jump back, which may give the prey a chance to escape. Here, the effect seems to be a kind of mimicry of the sudden nasty shock that a predator's own predator might give it.
Aposematism or warning colour. The function of warning colour is to deter prey from attacking prey that have active means of defence.
Warning colours are usually (but not always) bright and conspicuous. As in road-signs, bright, contrasty warning patterns are probably easier to learn: they are advertising colours. In practice, brightly coloured animals known to be defended, such as wasps and unpalatable Heliconius butterflies, are assumed to be aposematic - little experimental evidence shows they are actually easier to learn.
Learning a warning colour, to a first approximation, probably requires that the per capita loss during learning decreases as warning colours increase within a population. After predators experience unpleasantness with a warning pattern, they will learn avoid other individuals with the same colour pattern, whatever their density. So the fraction taken declines with the density of the warning coloured prey. So warning colours have a frequency-dependent fitness advantage that increases as the frequency of warning colours increases. Polymorphisms are therefore not expected within populations of warningly-coloured species, and indeed are rarely observed.
Once the majority of a population of predators knows the warning pattern, crypsis should become disfavoured, so it no longer pays to hide.
Warningly coloured species have to warn of some sort of punishment meted out on the predators that attack them (unless, of course they are fakes, i.e. mimics of another species - see below). Punishment could be in the form of:
Unpalatability including various forms of noxiousness (like wasp stings or irritating caterpillar hairs, unpalatability in the strict sense, or foul smell -- e.g. skunks. Distasteful and unpalatable species often obtain their poisonous compounds from plants (such as alkaloids, or glycosides that release cyanide when tissue is damaged), or they can synthesise the compounds themselves.Mimicry. A mimic evolves to look like an unpleasant model species. Two major kinds of mimicry are:Dangerousness is a common feature of warning coloured vertebrates, such as lethal coral snakes (cobra family). Warning colours are usually learned by predators. However, it has been shown that a reptile predator, a Mexican bird called the mot-mot, can innately recognize and avoid coral snake patterns. For coral snakes, this may be necessary because a mistake can mean death.
Batesian mimicry - after its discoverer Henry Walter Bates (1862). A Batesian mimic cheats; it is a parasite on the model's signal because it is itself palatable and pleasant to eat. Batesian mimicry hurts the model when the frequency of mimics becomes high, and so should have a frequency- dependent advantage that declines as the frequency of mimics increases. This probably explains the polymorphisms in Batesian mimics such as Papilio dardanus, an African swallowtail whose females mimic a variety of unpalatable models from the Danaidae: survival is higher is each mimetic form is rare relative to its model. The advantage is probably greater for the females, because males do not show the mimetic patterns; sexual selection probably helps to maintain this strong sexual dimorphism.Some examples of mimicry are [given in overheads]:Müllerian mimicry - after Müller (1879). A Müllerian mimic is itself an unpleasant species, and therefore should be a mutualist both with the model species and with the predator, whom it informs honestly of its unpalatability status. Müllerian mimicry works because, while a naive predator learns the pattern, it causes some deaths or injuries to the bearers of that pattern. Each predator will probably damage a constant number of individual prey during learning, so the more individuals there are with that pattern, the lower the per capita damage rate for the pattern. A Müllerian mimic has a frequency- dependent advantage that increases as the frequency of mimics increases, the opposite to Batesian mimicry. Müllerian mimicry is simply a multi-species generalization of warning colours within a species: Müllerian mimicry works within as well as between species. Müllerian mimics are almost always not sexually dimorphic; presumably because the mimetic advantage for increased frequency in these species outweighs any sexually-selected advantage of non-mimetic patterns.
A mimicry ring is a group of species all mimicking the same pattern. A ring usually consists of a backbone of abundant Müllerian mimics (or models, since each species acts both as a mimic and a model), but may also include Batesian mimics.
1) Butterfly mimicry rings.
1A) Butterfly Batesian mimicry rings with female-limited polymorphism.
1B) Müllerian mimicry rings in butterflies (e.g. Heliconius)
2) A vertebrate mimicry ring: coral snake mimicry.
Further examples of mimicry which are fun because they do not involve typical Batesian or Müllerian mimicry:Enough natural history!3) Cleaner fish, another vertebrate mimicry ring.
4) Cuckoo egg-mimicry. Yet another vertebrate example.
5) Orchid flowers mimicking Hymenoptera.
6) "Back-to-front" butterflies.
7) Egg mimicry in Passiflora
There is good experimental evidence for the efficacy of warning colour and mimicry.
Jane van Zandt Brower & Lincoln Brower in the 1950s performed laboratory experiments with blue jays and monarch butterflies (Danaus plexippus):
1) Jays fed monarchs became sick & wiped beakThere have also been several field experiments which successfully demonstrated mimicry. My favourite is by Jeffords, Waldbauer & Sternberg who worked on Batesian mimicry. They painted palatable male day flying moths (Callosamia promethea) to resemble unpalatable and palatable species. The saturniid moth they used was also a natural mimic of the unpalatable papilionid Battus philenor. They then recaptured the males by baiting traps with females, which "call" using pheromones. The recapture rate of mimics of palatable species was a mere 40% of that of mimics of unpalatable species; therefore Batesian mimicry was a very strong selective pressure, with a selection pressure, s=0.6. Similar field experiments with Heliconius (which have Müllerian mimicry) have shown similarly high selection pressures of s>>0.2-0.6. So selection for mimicry can be strong.monarchs are unpalatable.
2) Jays learn to avoid monarchs
3) Having learnt, Jays avoid mimics of monarchs
Evolution of unpalatability and warning colour
The problem. In the 1960s and 1970s when Hamilton's discovery of kin selection and sociobiology became generally accepted, it was realized that both unpalatability and warning colour could be seen as altruisms which could benefit the group more than the individual carrying them. This is the same problem as the evolution of alarm calls which may, like warning colour, attract the attention of the predator. Upon being eaten, a warningly coloured, unpalatable individual would be doing a great service to everyone else -- by teaching predators the errors of its ways -- but at great cost to itself! How could it evolve?
Altruistic behaviour, if it involves benefit to the population at the expense of costs to the individual should not evolve. Fisher in 1930 had suggested that unpalatability might have evolved because, although detrimental to the individual, it could be advantageous to the individual's siblings in groups of unpalatable caterpillars all laid as eggs by a single female. This was one of the first formulations of kin selection. Subsequently, in the 1960s -1980s, it became popular to ascribe the evolution both of warning colour and of mimicry to kin selection. But the arguments were simplistic, and didn't carefully consider the actual organisms and mechanisms involved; individual selection and/or the shifting balance could also be involved. Unpalatability and warning colour actually evolve in rather different ways. I here try to give a brief resumé of the arguments both for unpalatability and for warning colour.
Unpalatability is most like a true altruism of the two. The cost of this altruism consists of (a) metabolic energy needed to sequester or synthesize the unpalatable compounds, and (b) danger in teaching predators. But the danger in teaching is a probabilistic danger, probably roughly equal in all individuals bearing the compounds, so it cannot really be said to be a cost; if it is beneficial to the group, it also has a net probabilistic benefit to the individual also. The cost in sequestering compounds, itself may not be great; detoxification and excretion could cost more than putting the compound by in an inaccessible part of the cuticle.
On the other hand, the benefit to the individual itself of being unpalatable could be enormous; for example if compounds are sequestered in tough, leathery tissues that are prone to attack but not damage (such as the wings of Danainae, Acrainae, or Ithomiinae). It is now known that many birds taste-reject unpalatable butterflies relatively unharmed.
If the cost due to sequestration outweighs the individual benefit, unpalatability would be a true altruism, in that a warningly coloured cheat would do rather well by avoiding the cost of chemical sequestration, but gaining protection due to others' unpalatability. This is like, for instance, helping at mother's nest, if helping is costly in terms of ones' own reproduction.Living gregariously. So why do some unpalatable caterpillars live in family groups? Surely this is good evidence for kin selection! Nope! This just shows how wrong one can be! The problem is that there is a very good selfish reason why unpalatable species should live in groups (and they needn't be family groups). If prey can satiate their predators' appetites, it pays them to aggregate as much as possible. Most palatable, cryptic species cannot easily satiate their predators, so they often live solitarily, unless they are very abundant. Unpalatable species have a big advantage: they are specifically designed to swamp their predators. If instead of living scattered around where many different individual predators will find them, they can all crowd as a selfish herd into a small area where they gain an enormous per-capita advantage by quickly satiating the local predators, while avoiding other predators' territories altogether.If, on the other hand, benefits to the individual outweigh the costs, unpalatability won't be an altruism and will evolve under individual selection alone. It could be helped along by kin selection, if relatives are all close by, as in the caterpillars observed by Fisher. But this would not necessarily be the primary reason why unpalatability evolved. Individual benefit would explain why relatively solitary larvae, like the famous monarch butterfly Danaus plexippus and many Heliconius are unpalatable. Almost all the monarchs (Danaidae) are highly unpalatable, yet they all have solitary larvae, and spend their whole time as adults migrating hundreds, even thousands of kilometres, as though to be as far away from their relatives as possible.
So unpalatable species should be especially commonly found living gregariously. They are indeed often found gregariously, as larvae, usually (but not always) because they were originally laid as a single clutch of eggs by a single female. But they are also often found in aggregations as adults. Unpalatable Heliconius butterflies roost gregariously in unrelated groups of 2-50 individuals, often even with members of other species in the same Müllerian mimicry ring. North American Danaus butterflies fly south to Mexico in winter, and roost in enormous overwintering aggregations of tens of millions of individuals.
Kin selection may help in the evolution of unpalatability, as Fisher suggested, but aggregations are not good evidence that it is at all necessary! Aggregating behaviour of unpalatable species probably evolved after the evolution of unpalatability.
Warning colour has rather different evolutionary dynamics, as we have already mentioned. Our species will be assumed already unpalatable due to a sting, or to the sequestration of unpalatable chemistry from a host plant. There may be costs due to the production of warning colours, though we can reasonably assume that bright colours are about as cheap as the browns and greens of camouflaged species -- very different from the assumed costs of sequestration of nasty compounds.
Then, as in unpalatability, there are costs due to teaching predators. These are peculiarly frequency-dependent. When a warning colour pattern element first arises in an unpalatable prey, it should almost always be disfavoured. First, it is more conspicuous (to teach better). Second, no predators in the neighbourhood will have encountered the new pattern, so they will be naive; they will remember the old pattern somewhat, even if not very conspicuous or memorable.
Suppose the new pattern gets commoner, a strange thing happens. It becomes better to have the new pattern than to have the older, now rarer pattern: the newer pattern is now the pattern that predators do recognize (after a bit of learning, that is). Thus we have a special kind of frequency-dependent selection against rare forms. Whereas it is possible to interpret a newly evolved warning pattern as an altruism, a common warning pattern is hardly an altruism, because it pays to have it. When the trait is common, it would NOT pay to cheat. This is very different from helping at the nest, in which benefits and costs, at least within a family of specified relationship do not depend on the population frequency. It is also different from unpalatability, where, if it is altruistic, cheating may pay at high population frequency as we have seen.
It is therefore simpler to think of warning colours as a frequency-dependent trait with a disadvantage to rarity, rather than to think in terms of altruisms. The difficulty for the evolution of warning colour pattern is that selection is conservative, and acts against the novel pattern, even if it is a better warning signal. For example, below is a simple one-locus frequency-dependent fitness function: we assume that frequency dependent selection is linearly related to the frequency, with coefficients s and t (actual warning colour will have a more complex, non-linear function!).
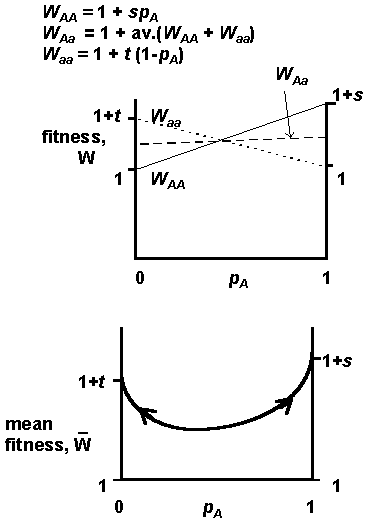
So how do novel warning colours evolve? If they are always selected against when rare, it is hard to imagine how a new warning colour evolves; whether in a cryptic species or an already warning coloured one. Although not exactly an example of a true altruism, a kin selection model (or at least an example of selection acting in groups of kin) may work, and was proposed in the 1970s. If a mutant phenotype A exists, it is more likely to be present in close kin than elsewhere. The new pattern might evolve locally, in groups of close kin, and then spread out to other groups.
If you are quick, you will have noticed that this "kin-selection" model is just a special case of the shifting balance. The first mutant will usually lack any family members with the new pattern, unless it is lucky to be a mutation early on in the mother's germ line. Normally, to have several local family members with the same allele, the mutant has become locally common, i.e. has drifted to a frequency considerably above zero, as in Phase I of the shifting balance. There are subtle differences, but essentially Phase II is now necessary, where selection increases the pattern locally provided it has crossed the adaptive trough, or threshold frequency. In Phase III, the new pattern will expand its range if fitter, either because it allows a greater population size and therefore causes emigration; or behind a moving cline.
Warning colours can also evolve by individual selection. You will have noticed that many palatable butterflies are already brightly coloured (peacock, red admiral in your garden). This can be because they signal to each other for sex - in fact butterflies formed a large section in Darwin's book on sexual selection. Or they might signal to predators, via flash coloration, or because they are Batesian mimics. If these brightly coloured butterflies were to suddenly become unpalatable, perhaps because of a switch to a new host plant, they would be preadapted to warning coloration. Alternatively, a newly unpalatable species might evolve warning colour very simply, by evolving Müllerian mimicry. Finally, any of these warning patterns might become enhanced by sensory bias, exactly as in sexual selection. For example, a flash-coloured moth might develop colours similar to the flash colours on the dorsal surface of the forewings, as well as the hidden hindwings, so enhancing the pattern.
Evidence
Most warningly coloured species belong to whole families that are unpalatable and warningly coloured, so that it will probably be impossible to work out exactly how the first species of the group, now probably extinct, evolved warning coloration. But a noticeable thing about warningly coloured clades is the diversity of warning patterns they present. We have already talked about warningly coloured races within species - these are very common; clearly, the evolution of warning colour is happening very rapidly, all the time, in spite of the evolutionary hurdles that we see in its way.
The very large Müllerian mimicry rings to which many of these species belong shows clearly, that the majority of species evolve warning colours by means of Müllerian mimicry. However, the sheer diversity of colour patterns is very hard to explain solely by a deterministic process such as mimicry, which by definition reduces pattern diversity. Instead, a minority of species, the models, must undergo enough random divergence, perhaps triggered by the shifting balance (at least according to Mallet!), to keep replenishing the diversity.
Warning colour,
mimicry, and speciation.
Bates,
Wallace and Darwin all thought that warning colour and mimicry were examples
of how natural selection might lead to the separation of a single species
into a number of different species. Today, mimicry is rarely mentioned
in the context of speciation. However, because selection is "intrinsic",
in the sense that it depends more strongly on the local frequencies of
colour pattern than on the abiotic environment, mimicry will contribute
to speciation by selecting against intermediates that lie between two adaptive
peaks. This will be similar to other intrinsically selected traits such
as such as chromosomal rearrangements and genes under epistatic selection.
We know that selection pressures on warning colour and mimicry are intense,
and that many of the same principles apply to all these traits. Closely
related aposematic species often differ widely in their warning patterns,
paralleling the case for chromosomal rearrangements. Obviously, not all
speciation is via warning colour, but warning colour and mimicry are good
external examples of exactly those traits which normally have their hidden
effects deep within the reproductive system, and do generally cause
speciation.
We here cover some evolutionary oddities about the evolution of mimicry. Goldschmidt in 1940-1945, proposed that, while gradualistic, Fisherian multilocus evolution could explain evolution within species, speciation was due to a different process that he called "systemic mutation" - single, massive chromosomal mutations that reorganized the whole genome. But he suggested that there were also some examples of systemic mutations within species; firstly chromosomal mutations, and secondly in mimicry. He knew that some results on Batesian mimicry showed that single loci apparently radically affected whole colour pattern, and even wing shape traits. Goldschmidt argued that these single genes were systemic mutations using pre-existing developmental genetic pathways present in the model to cause accurate resemblance in the mimic. In other words, the mimic obtained mimicry without selection, it was a "hopeful monster" whose adaptation was created by a single mutation then preserved by selection, but not constructed by microevolutionary multi-locus selection.
Fisher and his followers took up the challenge, and many experiments were performed on mimetic butterflies in the 1960s until the present day to attempt to refute these heretical ideas.
Here, we examine the evolution of mimicry
in the light of what is known about the genetics of mimicry, and try to
assess how far Goldschmidt was correct.
Much of the work on the evolution of Batesian mimicry was done by Sir Cyril Clarke and Philip Sheppard, on Papilionidae.
It was quickly realized that Goldschmidt was wrong on the details. Some of the best examples come from the Asian Batesian mimic Papilio memnon, whose polymorphic females mimic a variety of unpalatable model species, also within the Papilionidae.
1) Many features of mimicry showed evidence, for clearly analogous resemblances rather than developmental homology. For example, in the overhead, the female forms of P. memnon have white or red "shoulder flashes" on the wing base. These flashes mimic similar marks on the thorax of the model. Although these marks are mimetic, they are clearly not developmentally homologous.2) Mimicry is not inherited entirely as a single gene, as Goldschmidt's single mutation hypothesis requires. There are loci with major effect, but there are unlinked components also. When the single major locus in Papilio memnon was carefully analysed, extremely rare recombinants within the supergene were discovered both in laboratory broods, and as rare variants in the wild. Clarke and Sheppard were able to show that the apparent "single locus" developmental switch was in fact a "supergene" composed of a number of tightly linked elements. The recombining elements could be mapped in order in the manner shown on the overhead. Clearly, then, selection had built up the mimicry "bit by bit", and not via a single mutation.
3) Polymorphism can only be preserved in Batesian mimicry if tight linkage disequilibrium (enforced by the tight linkage) between these epistatic elements is preserved, so that non-mimetic intermediates are very rare. Instead of the single gene nature of the switch locus being evidence for a single locus, we can see that the mimetic polymorphism can ONLY evolve if the genes that are involved exist within a tightly linked section, i.e. a supergene. Here is a selective explanation for the very data that Goldschmidt used as evidence for his systemic mutation theory in mimicry. [MORE]
In Müllerian mimicry, we expect few polymorphisms, because selection acts against rare forms - this is, after all, the rationale for Müllerian mimicry. In general, this is true: lthough Müllerian mimics have much geographic diversity, implying rapid evolution of new warning colour patterns; within any one area, there are few polymorphisms. The major exception is in the narrow hybrid zones between races.
Therefore, the arguments applied to Batesian mimicry, that patterns should be inherited at single locus "supergenes", do not apply. In Heliconius, a Müllerian mimic, different geographic races have very different colour patterns (see top of this web page: H. erato (above) and H. melpomene (below)). A number of unlinked loci affect colour pattern inheritance in the Müllerian mimics.
On the other hand, it is clear that mimicry is hardly based on multiple unlinked genes of individually small effect, as suggested by Fisher in his theory of evolutionary adaptation. In this species, the major pattern changes is inherited at relatively few genes (or tightly linked gene families). Are such major gene effects a result of developmental genetic constraints? Or is there a selective reason why this might be so?
Sheppard and Turner have proposed a selective explanation which seems likely. A small mimetic change away from the mean for each species A or B will be selected against, since the protection will be reduced. Only a relatively major mutation will bridge the gap. If species A is commoner and/or more heavily protected, it can never be selected to evolve towards the more weakly protected species B. Only B will evolve, and then towards A.
Thus we again have a selective reason why,
even in Müllerian mimicry, a rather radical mutation may be favoured
over slow, polygenic adaptation. The reason is analogous to the case of
the shifting balance (which, incidentally, could provide another route
to Müllerian mimicry). Mimicry provides a "rugged" adaptive landscape,
with some high adaptive peaks among the possible phenotypes, but many adaptive
troughs or lowlands. A major mutation, a diminutive "hopeful monster",
if you like, does help Müllerian adaptation. On the other hand, once
near the adaptive peak, climbing the peak is done in the ordinary Fisherian
mode, in small steps via polygenic adaptation.
In summary, mimicry is a great example of evolution by natural selection, and has been celebrated for this ever since Darwin first wrote a glowing review of Bates' paper in 1863. But there are still many puzzling aspects of mimicry that need work. In particular, it would be nice to know a little about the molecular nature of the genes involved, and whether random drift may initiate some of these changes.
FURTHER READING
General textbooks. There are also some good specialist books on mimicry; you might try books by Wickler or Edmunds. You could also try the following (a shameless plug for my own work!):
Joron, M. & Mallet, J. (1998). Diversity in mimicry: paradox or paradigm? Trends in Ecology and Evolution 13: 461-466. . REPRINT - some pictures, recent controversies
Mallet, J. and Joron, M.
(1999). Evolution of diversity in warning colour and mimicry: polymorphisms,
shifting balance and speciation. Annual Review of Ecology and Systematics
30:201–233. REPRINT
- rather long!