 Close
Close

Structural and Molecular Biology
We are the Research Department of discoveries, new ideas and methodologies in biomolecular sciences. Within the stimulating and diverse research and training environment of international standing, we lead and advance on essential questions in exciting areas of modern biochemistry, biotechnology, and medicine at the atomic, molecular, cellular and organism levels.
Owing to the strength and the technical expertise of its research community SMB is a key player in many collaborative multi-disciplinary projects across UCL, nationally and internationally. This vision is enhanced via our partnership with the Department of Biological Sciences at Birkbeck College and the Institute of Structural and Molecular Biology (ISMB).
Emerging from the rich history of Biochemistry Departments at UCL, nowadays, the SMB provides a home to engaged, enthusiastic and well-supported staff and students. We learn and teach through our research; we nurture curiosity, welcome challenging questions, and embrace all voices.
 462:2011-2015)")
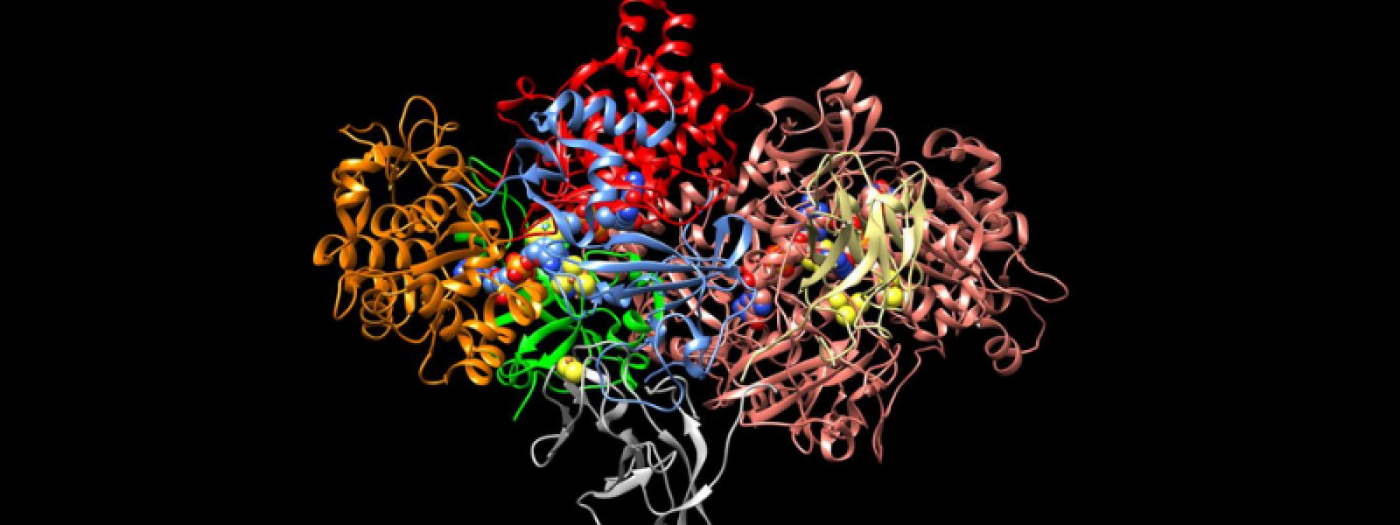
Atomic resolution structure of the core complex of a type IV secretion system involved in bacterial conjugation (Nature (2009) 462:2011-2015)
Our Department



Research
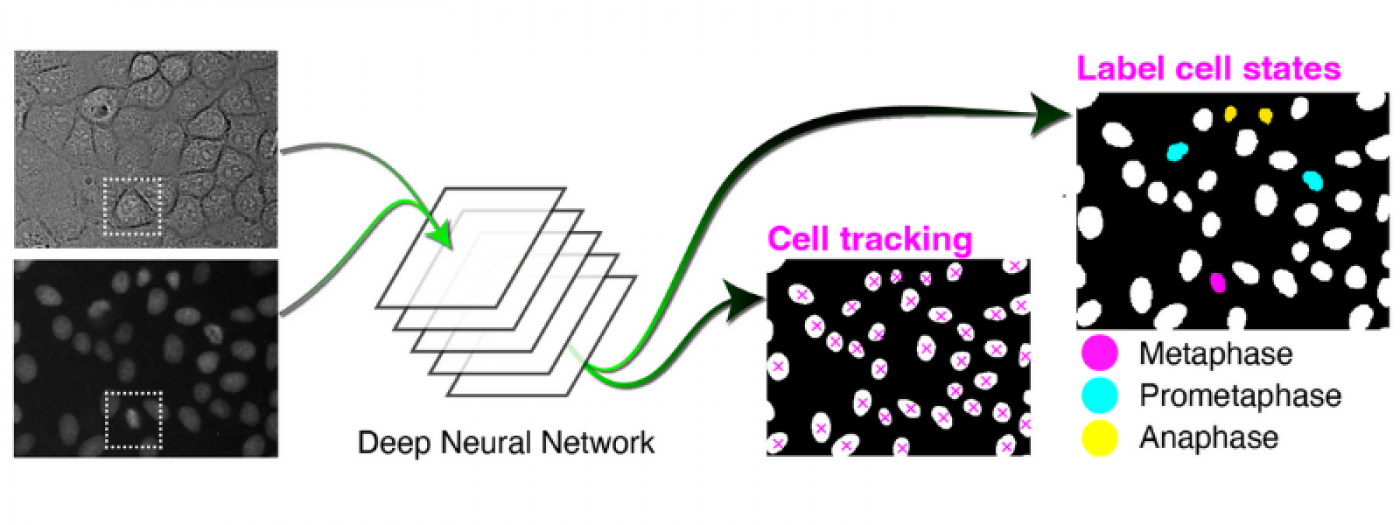
Computational Biology and AI
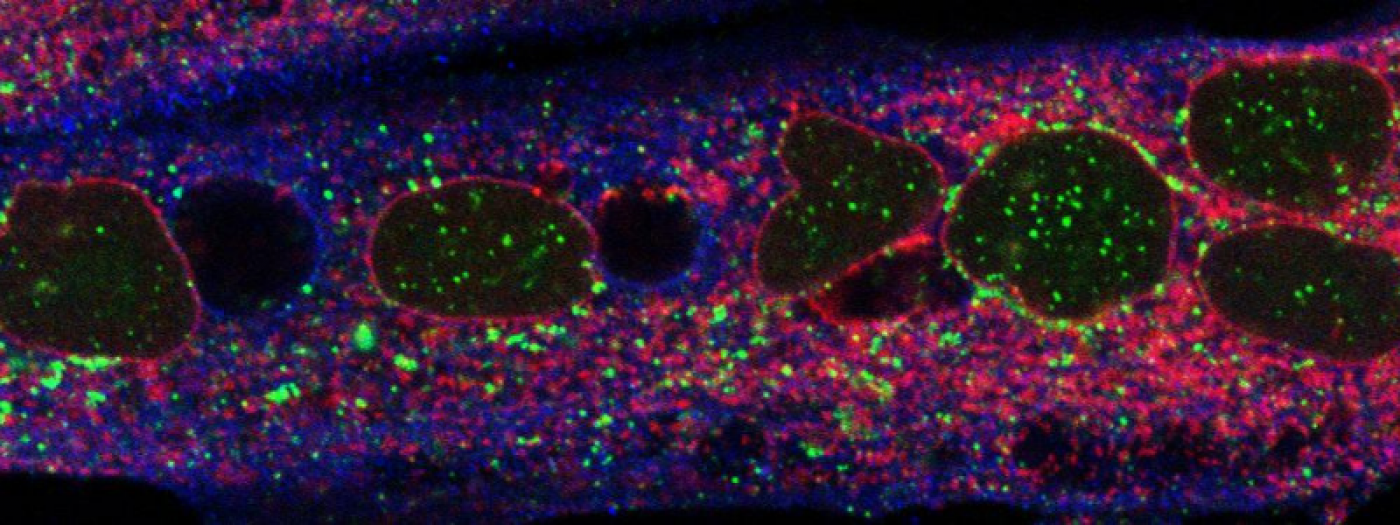
Membranes, Signalling and Trafficking
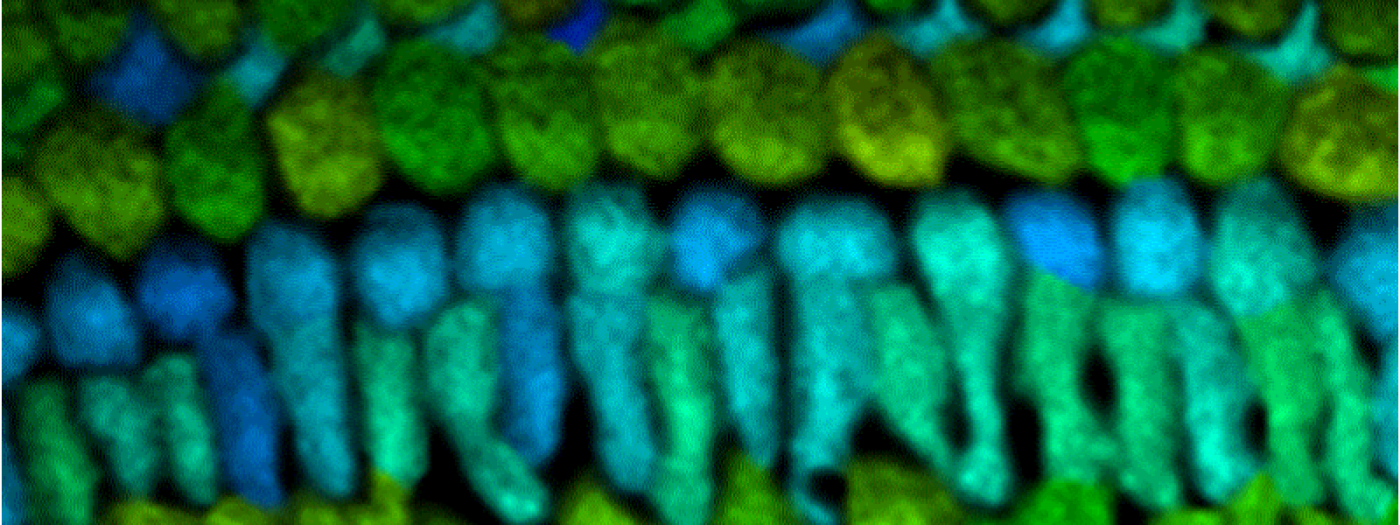
Metabolism and Disease
Microbes and Environment
Protein folding in health and disease
Transcription and RNA Biology

Research Groups
Find out about the ground-breaking research work that is going on across the 30+ research groups which make up our department.
Research Themes
SMB develops innovative research in modern biochemistry, biotechnology, and medicine at the atomic, molecular, cellular and organism levels through six research themes.
Early Careers Community
Our research department is home to a thriving community of postdoctoral researchers and fellows supported by skills training, careers advice and more.
Fellowships
We promote multi-disciplinary research at the interface of structural, cell and chemical biology in a unique link between UCL and Birkbeck and welcome fellows.

The Department hosts a number of facilities that underlie our work in the biophysics, molecular structure, biochemistry and cell biology fields.
SMB on Youtube
Watch our researchers presenting their work at conferences and events all over the world.
Study
We offer BSc and MSci Degrees in Biochemistry. Students in Biochemistry or Biological Sciences are also able to enter a Molecular Biology pathway in Year 2.
Biosciences offers a range of MSc and MRes programmes. In particular, the MRes Biosciences offers 12 pathways including Biochemistry.
The department offers both three and four year MPhil/PhD courses covering a variety of research projects.
Short Courses
Short and intense training for anyone wishing to learn new skills or further develop the ones they have. Courses have a high tutor/student ratio.

SMB 3 Minute Thesis Winner
Is it possible to present an 80,000 word thesis in 3 minutes in non-specialist language with just one slide?

Institute for Structural and Molecular Biology
The Institute for Structural and Molecular Biology (ISMB) is a joint initiative between UCL and Birkbeck that promotes multi-disciplinary research at the interface of structural, computational and chemical biology.


Find resources and support for promoting student and staff wellbeing

Find out about sustainability initiatives for labs and individuals
Contact Us
Department of Structural and Molecular Biology
Division of Biosciences
UCL Darwin Building
Gower Street
London WC1E 6BT
General Enquiries tel: +44 (0)20 7679 2308, email: tabitha.owen@ucl.ac.uk
Postgraduate Enquiries email: biosciences.pgr.admin@ucl.ac.uk
Undergraduate Enquiries email: biosciences-admissions@ucl.ac.uk
The majority of our laboratories and research spaces are located in the UCL Darwin Building on Gower Street, formerly the site of 'Macaw Cottage', a residence of Charles Darwin's.
The main reception to the building is accessed via Darwin Walk, off Malet Place, entered via Torrington Road.
UCL Map for Darwin Building entrance