Chondrichthyes
Chondrichthyes - cartilaginous fishes
| Vertebrata; Gnathostomata; Chondrichthyes |

Torpedo ray - dorsal view
|

Torpedo ray - ventral view showing toothplate
|
Torpedo ray - ventral close-up showing pelvic claspers
|
Diversity and Lower Taxonomy:
- Subclass Elasmobranchii (sharks,
rays and skates)
- Superorder Batoidea (over 500 species of rays and skates):
- Order Rajiformes (common rays and skates)
- Order Pristiformes (Sawfish)
- Order Torpediniformes (electric rays)
- Order Rajiformes (common rays and skates)
- Superorder Squalomorphii:
- Order Hexanchiformes: containing 5 extant species
within 2 families, the Hexanchidae(cow shark)
and Chlamydoselachidae (frilled shark), distinguished by their additional
gill slits (either six or seven).
- Order Squaliformes: containing 80 species divided into 3
families: Echinorhinidae(Bramble
Sharks), Squalidae (Dogfish sharks) and Oxynotidae (Roughsharks).
- Order Pristiophoriformes: containing
5 species within a single family,Pristiophoridae(sawsharks).
- Order Hexanchiformes: containing 5 extant species
within 2 families, the Hexanchidae(cow shark)
and Chlamydoselachidae (frilled shark), distinguished by their additional
gill slits (either six or seven).
- Superorder Squatinomorphii:
- Order Squatiniformes: containing 13 species in 1 family, Squatinidae (Angel sharks).
- Order Squatiniformes: containing 13 species in 1 family, Squatinidae (Angel sharks).
- Superorder Galeomorphii:
- Order Heterodontiformes: 8 species
in 1 family, Heterodontidae (bullsharks).
- Order Orectolobiformes: 7 families
are found within this order: Brachaeluridae (blind catsharks and blind sharks), Ginglymostomatidae (nurse sharks), Hemiscylliidae(bamboo
sharks and longtailed carpetsharks), Orectolobidae (wobbegongs),Parascyllidae (collared carpetsharks), Rhincodontidae and Stegostomatidae(zebra
sharks).
- Order Carcharhiniformes: this is
the largest order containing approximately 200 species within 8
families: Carcharhinidae (requiem sharks), Hemigaleidae (weasel sharks),Leptochariidae(barbeled
houndsharks), Proscylliidae (finback catsharks), Pseudotriakidae(false cat
sharks),Scyliorhinidae (cat
sharks), Sphyrnidae (bonnethead sharks, hammerhead
sharks, and scoophead sharks), Triakidae (houndsharks, smooth-hounds, topes,
and whiskery sharks).
- Order Lamniformes: known as
mackerel sharks, this order contains 7 families and 16 species:Alopiidae (thresher sharks), Cetorhinidae (basking sharks), Lamnidae (mackerel sharks, porbeagles, and
white sharks), Megachasmidae (megamouth sharks),Mitsukurinidae (goblin sharks), Odontaspididae (goblin sharks, sand sharks, and
sand tiger sharks) and Pseudocarchariidae (crocodile sharks)
- Order Heterodontiformes: 8 species
in 1 family, Heterodontidae (bullsharks).
- Superorder Batoidea (over 500 species of rays and skates):
- Subclass Holocephali (chimaeras)
- Order Chimaeriformes: 40 species
in 6 genera and three families, Callorhinchidae,Chimaeridae and Rhinochimaeridae.
- Order Chimaeriformes: 40 species
in 6 genera and three families, Callorhinchidae,Chimaeridae and Rhinochimaeridae.
 sensu Maisey (1980)")
This view was strongly contested in the following decade,
with many morphological phylogenetic studies (Shirai, 1992, 1996; de Carvalho,
1996) gathering support for the hypnosqualean hypothesis, which states that
batoids (rays and skates) are in fact derived sharks, grouped with the
Pristiophoriformes and Squatinomorphii in the clade "Hypnosqualea"
(see cladogram below). The remaining squalomorph orders (Squaliformes and
Hexanchiformes) then grouped (whether as a clade or grade) nearest to the
Hypnosqualea, to form the "Squalea", which formed the sister group to
the remaining elasmobranchs - the monophyletic Galeomorphii. In addition to the
controversial proposal that sharks are in fact paraphyletic with respect to
rays and skates, these studies were significant in that they began to identify
the fragility of the superorder Squalomorphii as a natural grouping, in these
instances containing both the batoids and the squatinomorphs. This, over time,
has led to the break down of the Squatinomorphii as a distinct elasmobranch
superorder, with its single order (Squatiniformes) being redefined as an order
within a newly defined Squalomorphii (containing its original three orders plus
Squatiniformes).
 sensu Shirai (1992, 1996)")
Since the turn of the millenium, a wealth of molecular
studies starting with Douady et al (2003) have brought about a new view of
elasmobranch phylogeny that strongly and
consistently rejects the hypnosqualean hypothesis, and returns to the
classically held view that sharks form a monophyletic group, the
Selachimorpha, which is sister to the rays and skates (Batoidea). Within
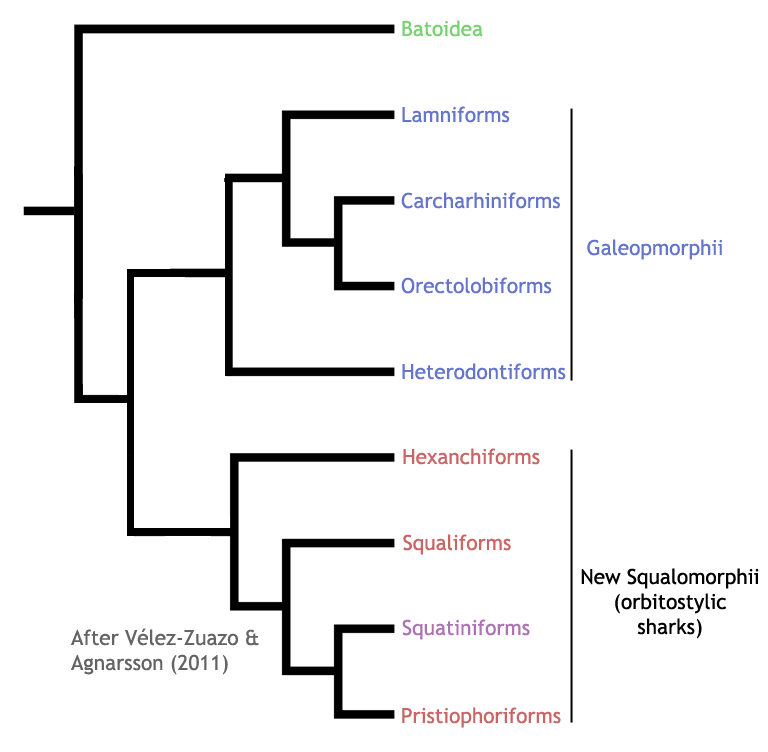
Selachimorphi, there has been repeated support for the monophyly of the new Squalomorphii
containing squatiniform sharks (Douady et al, 2003; Winchell et al, 2004;
Naylor et al, 2005; Mallatt et al, 2007; Heinicke et al, 2009; Vélez-Zuazo
& Agnarsson, 2011). Note that this newly defined Squalomorphii is
equivalent in terms of membership to the orbitostylic sharks. The phylogenetic
positions of the remaining four orders (galeomorph sharks) have, in contrast,
been less consistent. While some studies have recovered amonophyletic
Galeomorphii (Heinicke et al, 2009; Vélez-Zuazo & Agnarsson, 2011), others
have placed the Heterodontiformes as the closest living relative to the
Squalomorphii (Winchell et al, 2004; Mallatt et al, 2007). Most molecular
studies have, nonetheless, recovered the remaining galeomorph orders
(Orectolobiformes, Carcharhiniformes, and Lamniformes) as a monophyletic
grouping (see Douady et al, 2003 for exception), although the interelationships
between these groups is still a matter of debate. The two cladograms below show
recent conflicting views on the phylogeny of Selachimorpha based on DNA
sequence data.
 sensu Mallatt al (2007)")
Distribution and Habitat:
Most chondrichthyans are marine species. Only 5% (approximately 45 species) are restricted to freshwater, such as the giant freshwater stingray, Himantura chaophraya (Fowler et al., 2005; Helfman et al., 2009). Others enter estuaries and freshwater sporadically, often to breed.
Members of the Chondrichthyes can be found in nearly all aquatic ecosystems and depths, except the most extreme conditions. But most species are restricted to and specialised for a particular oceanic zone. For example, the skates (members of Rajidae) and angel sharks (Squatinidae) are benthic species. The Lamnidae (white sharks) are pelagic, but in the upper depths only (Fowler et al., 2005). Approximately 50% of species inhabit the continental slopes, up to 200m, and 35% occupy depths between 200 - 2000m (Helfman, 2009). Only 5% of species, mainly the large pelagic ones, inhabit the open ocean. For example, the mantarays (Myliobatiformes) make annual migrations, while the great white shark (Carcharodon carcharias) has been known to travel between South Africa and Australia (Fowler et al., 2005).
The elasmobranchs occcupy the widest range of habitats - even the icy waters of the Arctic and Antarctic Ocean. But their physiology cannot withstand the most extreme conditions such as high salinity or the oxygen deficient depths that some teleosts have become specialised to endure. Therefore most sharks do not inhabit very deep water, although some species have been sighted at depths up to 4000m (Helfman et al., 2009). The Holocephali are found only in temperate waters up to 2000m, and prefer the cooler depths below 80m (Fowler et al, 2005; Helfman et al., 2009).
Conservation Status (IUCN):
The 2008 IUCN Red List categorised 17% of chondrichthyan species (181/1044) as threatened with extinction. The majority of these are in the lowest risk category (Vulnerable), but 42 species are Endangered, and 25 are placed within the highest risk category (Critically Endangered). The Squatiniformes (angelsharks) are the chondrichthyan order at greatest risk, with over half (12/22) of its species considered threatened with extinction (3x Critically Endangered, 5x Endangered, and 4x Vulnerable). Seven of the remaining ten species are Data Deficient, and so may themselves also be under threat.
Chondrichthyes are an ancient and successful clade, having survived 400 million years of changing environments. Yet human presence and intervention has put them at risk. The life history of Chondrichthyes marks them as vulnerable to extinction. As a K-selection species, they give birth to only a few young after a long gestation, which grow slowly and reach sexual maturity late (Fowler et al., 2005). Although their rate of survival is usually high, if populations become rapidly depleted, recovery can be difficult and lengthy.
The primary threat to sharks, rays, and chimaeras is fishing (Fowler et al. 2005). Commercial demand for shark meat and fins is high and fisheries are unmanaged, pushing population levels to dangerously low numbers. Many species are also often killed as by-catch from bottom trawlers.
Loss of habitat is also a significant threat, and those species which inhabit regions closest to human activity are at most risk from habitat degradation and pollution. Estuaries, whose shallow waters provide invaluable nurseries for chondrichthyans are being destroyed or polluted. Similarly, certain freshwater systems are becoming increasingly degraded by pollution, making them inhospitable to indigenous species.
Features:
- Skeletons
formed of calcified cartilage - no
bone.
- Covered
in placoid scales - a structure like teeth, with a dentine crown coated in
an enamel-like material, a vascularised pulp cavity, and a bony base.
They are sometimes referred to as denticles.
- True
teeth, which are shed and replaced regularly in modern species.
- The
males of all but the oldest fossil species have a pelvic clasper, used in courtship
and mating. It is formed of the pelvic meta-pterygium - the basal cartilage
of the pelvic fin.
Licensed under the Creative Commons Attribution Non-commercial Share Alike 3.0 License
'Vertebrate Diversity' has been released as an open educational resource (OER) on a Creative Commons 'Attribution Non-commercial Share Alike' license. This means that once downloaded, content can be modified and improved to complement a particular course. This requires, however, that improvements are recycled back into the OER community. All content present at the time of download must be accordingly credited and, in turn, novel content must be appropriately licensed. For more information, please refer to the license deed by clicking on the link above.