Next lecture, we shall be talking about speciation. So here we need to cover topics about the nature of species, and discuss how and whether speciation is different from microevolution (evolution within species)?
- What are species?
- How do species differ from each other?
- How many species are there? We will
briefly cover species-level biodiversity.
Darwin in 1859 proved to the world (the reasonable part of it, anyway!) that species evolved, rather than were created. But this made for a difficulty. All of a sudden species weren’t created kinds, with an Aristotelian essence, as previously thought. It then became unclear how species differed, if at all, from other categories. Species evolve from non-species, so where is the dividing line? Darwin hard a hard time with this one, because if species didn’t exist, he could hardly write a book on their origin, could he?!
Darwin’s resolution of this conundrum was to use a pragmatic definition of species - sometimes dismissively called the morphological species concept, in which species were distinguished from races and polymorphic forms by drawing a suitable dividing line in the actual continuum between species and races or forms.
It would be nice to say that there the story ends. Unfortunately, it doesn’t. Species concepts have been for the last 10-20 years a major battleground for systematists, philosophers of biology, and evolutionists. My own view is that Darwin was thinking more clearly than many of the modern contestants, even though his theory of genetics was patently wrong. But not many agree with me (yet!). I will therefore attempt to give you a fairly balanced assessment.
So here are just some of the leading "species concepts", and their strengths and weaknesses.
1) The morphological species concept (phenetic species concept also included)
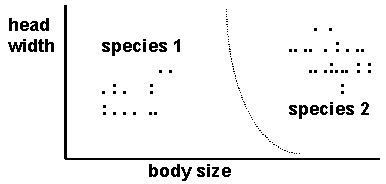
According to Darwin, species can simply be diagnosed by morphological gaps in the variation between individuals (see diagram above, where the line separates two morphological clusters of individuals). For instance, Darwin regarded Primula veris (the primrose) and Primula elatior (the cowslip) as varieties of the same species because many intermediates or hybrids are found between them. He argued in the same way that the many races of humans were members of the same species. In these cases, it is not easy to find a sensible place to put a dividing line, even though there are clear differences between the forms. Darwin’s ideas were revived by numerical taxonomists in the 1960s, who introduced a multivariate statistical version of the idea, known today as the phenetic species concept.
However, Darwin’s ideas do lead to some problems:
a) Variation within species sometimes leads to morphological gaps. For instance, we have seen that races, subspecies, populations and even morphs within populations are often discrete (i.e. the variation is discontinuous, there are gaps). Nowadays, we would certainly not classify the melanic form of the peppered moth as a different species just because the variation is not continuous.
b) Lack of differences between species: There are oftensibling species which (a) are morphologically more or less identical, although genetically different, (b) evolve more or less separately, (c) have little or no hybridization or gene flow between them. Some examples are:
2) The biological species concept
Difficulties with Darwin's concept tempted a number of people to try to redefine species by means of interbreeding. These ideas were first put forward clearly by an entomologist, E.B. Poulton in 1903. Later, Dobzhansky (1937), and, most famously, Mayr (1940, 1942, 1954, 1963, 1970 etc. etc.) carried on and popularized this tradition; it was Mayr who named the idea the "biological species concept", thereby unfairly trying to take the high moral ground because anyone else’s species concept was thereafter, of course, "non-biological"!
The biological species concept allows for abundant gene flow within each species, but a lack of hybridization or gene flow betwen species. The lack of gene flow is caused by isolating mechanisms, a term invented by Dobzhansky, but again popularized by Mayr. Because they are not necessarily "mechanisms" in any sense, I prefer the term "reproductive isolation":
Types of reproductive
isolation
A) Pre-mating
isolation {or
pre-zygotic
isolation}
a) Ecological or seasonal isolation -
mates do not meet
b) Behavioural (biochemical) isolation
- individuals meet but do not attempt mating
c) Mechanical isolation - attempts at
mating do not work!
B) Post-mating
{orpost-zygotic}
isolation
d) Gametic incompatibility - gametes die
before fertilization } (note: this is post-mating but pre-zygotic)
e) Hybrid inviability - hybrids have reduced
viabilility as zygote or later in development. This may be caused by internal
(genomic factors), or because hybrids are not suited to survival for ecological
reasons. Hybrids may also have reduced mating propensity, or be disfavoured
as mates.
f) Hybrid sterility - hybrids survive
and mate as normal, but are partially or completely sterile.
g) Sexual selection against hybrids (studied
by Russ Naisbit, a PhD student in my laboratory) - hybrids are healthy
and fertile, but disfavoured during mating.
Problems with
the biological species concept
a) Does not apply in allopatry.
Strictly, the biological species concept only works in sympatry and parapatry,
because how can we tell whether two species would intercross if they are
allopatric? We can put them together to see if they interbreed, but many
sympatric species of ducks, Drosophila, even tigers and lions will
interbreed in captivity, though they rarely, if ever, do in the wild.
So when two species are allopatric, we have to guess from their traits - morphology, behaviour, genetics - and their behaviour in captivity, if possible, whether they would interbreed if they were in sympatry. Not very scientific? (This is a problem with all definitions of species that propose a single fundamental essence of species. Given that species originate by evolution, species identity is bound to be more dubious the more time that they have been diverging. Thus species are bound to become less real and more difficult to classify with increasing spans of space (in geography) or time (in the fossil record).
b) Natural hybridization and gene flow between species exists.
Around 10% of birds and butterfly species produce hybrids in the wild, although each species usually does so very rarely (maybe 1/1000 or less). Ducks [SEE OVERHEAD] and other birds of paradise seem particularly prone to hybridization (>50% of species) in the wild, even though most of the time they seem like "good" species. Fewer mammals probably hybridize; in Europe, only about 6% of species are known to form hybrids in the wild. However one of these is the world’s biggest animal (ever - beats the dinosaurs hands down): the blue whale, has been recorded hybridizing with its near relative, the fin whale. Not only that, a female hybrid between these two species has been found with a healthy foetus, genetically a backcross. Plants are especially well known for their tendency to hybridize (probably well over 20%), and hybridization is even a major source of speciation by allopolyploidy in this group (see below). For this reason, the biological species concept has never really caught on with botanists.
Hybridization would not matter if genes did not pass between species via hybridization. But we now know that genes DO pass between species, and many species have received genes, or whole mitochondrial genomes from other species. In some cases, flowering plants have even adopted genes from symbiotic bacteria. DNA sequencing has now revealed many, many examples of this kind of horizontal gene transfer between species. Hybridization and gene transfer are today very important topics in conservation and economic biology.
Although the biological species concept has long been accepted by many evolutionary biologists (especially zoologists) as the best species concept, these kinds of problems have led to increasing attacks. Several possible solutions have been proposed.
3) Ecological species concept
Leigh Van Valen, in the 1970s suggested that species were better defined by the types of selection they underwent, or by their ecological niche. Real species, argued Van Valen, are ecologically different.
Problems
a) It is at least theoretically possible that some kinds of sibling species might have exactly the same niches. Eventually, this would lead to a probable loss of one of the species through competition, so this problem is perhaps more theoretical than actual.
b) The worst problem for this idea is that species often do have ecological morphs within the species. The cichlid fish Cichlasoma from Cuatro Cienagas, Mexico, has multiple morphs that do different things:
4) Cladistic and phylogenetic species concepts
Recently, most systematists have favoured phylogenetic systematics, in which cladistic classifications. The cladistic movement was founded by Willi Hennig in the 1950s. If higher taxa are defined by means of phylogeny, then so should species, reasoned cladists. This has led to a plethora of cladistic and phylogenetic species concepts. One idea, based on Willi Hennig’s own idea, and supported by Ridley among others is a cladistic species concept:
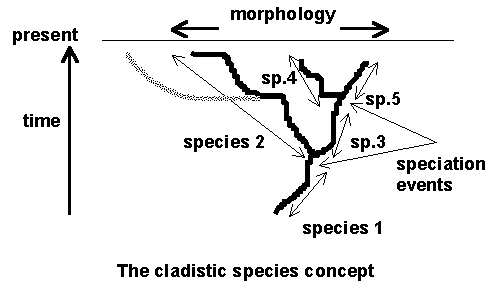
According to Hennig and Ridley, species are branches in a lineage. When the lineage branches, two new species arise out of the old one, as above, where 5 species result from a phylogeny with two branching (speciation) events. Although there is a morphological discontinuity within the history of species 2, this does not mean the upper and lower portion of species 2 are different species, unless a new branch (in grey) originates at that point. The virtue of this idea to its proponents is that it should apply in history, to fossils, as well as to modern species.
Unfortunately, there are Problems:
a) In practice, phylogenies are unstable hypotheses rather than facts. The branching pattern must be known in order to define species. Cladistic species may therefore be somewhat arbitrary. Supposing the grey branch was unknown, then was suddenly discovered in a small modern population. Now, suddenly, fossil species 2 must be reclassified into two separate species, even though a continuous record of those forms were previously well known. Worse, if the grey lineage has no fossil record, we don’t know where species 2 must be divided.
b) Many island populations may be cladistic side-branches of mainland species; yet their establishment does not usually alter the mainland species in any way whatsoever. In fact, this may be true for any population that has been geographically isolated for a few generations. Cladistic species concepts could lead to a lot of new species that are only faintly recognizable.
c) Hybridization, if it occurs between branches, will tend to lead to a lack of clear branching between related pairs of species at some genes. The phylogeny of species may be meaningless under such conditions; instead, the phylogeny becomes a mass of "genealogies" at sometimes contradictory genes. Of course one could use some sort of average phylogeny (sometimes called a "consensus" phylogeny) as the "true" species phylogeny, but this kind of averaging is certainly very different from the notion that the species we are looking at have a single true phylogeny.
There are many alternative evolutionary and phylogenetic species concepts which attempt to answer these problems. For example, various kinds of phylogenetic concept have attempted to incorporate the possibility of gene flow between species. For instance, Cracraft suggests that species have fixed differences at (morphological) "characters", but critics have argued that this would lead to the recognition of many local populations with trivial genetic differences as separate species. It is also a little unclear what one means by "fixed" differences when gene flow will prevent complete fixation. We don’t have time to go through all the species concepts of this sort here, but you can find them in many books (some of my own efforts, encyclopaedia entries with general references, are available from my home page).
5) Rank-free taxonomy, and giving up on species altogether!
Recently, a number of leading phylogenetic systematists have proposed "rank-free" taxonomy, in which species no longer hold a unique position in the taxonomic hierarchy. Proponents of this view argue that the difficulty of assigning a species rank exists because species lack reality as special taxa. Instead, they argue, we should develop a completely new taxonomy based purely on phylogenetic principles, and do away with the Linnean binomial (i.e. two Latin names: genus + species) tradition. The first revision of a taxonomic group without species designations has recently been published in the journal "Systematic Biology" (2000). Whether this idea will catch on is hard to say. If it does, it could cause chaos in biological nomenclature at a time when we badly need taxonomists for studies in biodiversity and conservation. There is a very strong resistance to this idea from among traditional taxonomists, and also from within even the phylogenetic systematists.
My own view is that hybridization and gene flow will wreck the idea of the perfectly hierarchical rank-free taxonomy, especially near the (current) species level, and that species will remain a convenient naming device to classify animals and plants. There must be a certain validity to species, or your bird or plant guides wouldn't be very useful. In some asexual taxa, like brambles and dandelions, it may be somewhat difficult to distinguish "species" from "varieties", but mostly even asexual taxa are easy to divide along species lines. On the other hand, I rather agree that the supposed "reality" of species over and above other higher (genera, families) or lower (subspecies, varieties) taxa has been greatly overemphasised.
Why are there so many species concepts?
What should practising evolutionary geneticists like you do, faced with such a diversity of opinion?
I have my own way of making sense of this debate, with which you may or may not agree. I argue that you can update Darwin’s idea of species without too much difficulty, but take account of modern knowledge of genetics, and thereby solve some of the problems inherent in the other species concepts at the same time.
Species within a region are genetically differentiated populations potentially connected by gene flow. This gene flow may be very low (as in the biological species concept), but it doesn’t have to be negligible. The important thing is that the gene flow is low enough, and the disruptive selection keeping the populations apart is strong enough so that genetic differences between the species are maintained. If the two populations collapse together, because the gene flow outweighs selection, then there will only be a single species.
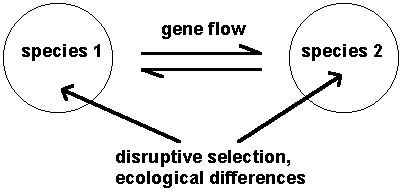
Species are then clusters of genotypes with discontinuities or gaps between them (a genetic version of Darwin’s morphological concept). Low levels of gene flow (a lack of Mayr’s pre-mating isolation) could break up the genotypic and phenotypic differences. However, this gene flow, if it exists, may be balanced by disruptive selection, which may be intrinsic (due to interactions between genes within the hybrids, as in Mayr’s post-mating isolating mechanisms) or extrinsic (due to the environment, as in Van Valen’s ecological concept). Darwin’s morphological concept can thus be related to the ecological and biological concepts: the biological and ecological concepts are explanations of the morphological/genotypic situation of two clusters separated by gaps. Phylogenies obviously have something to do with the whole process. As species diverge more and more, hybridization will be reduced, and a separate branch in the phylogeny emerges from the cross-linking caused by hybridization, and becomes progressively better defined.
Phew! Now that’s over, let’s get on with discussing the interesting things about species.
To study speciation, we need to know how species differ from one another genetically. In general, weherever we look, species differ in ways similar to those of populations or geographic races (see EVOLUTION IN SPACE AND TIME), only more so. Here are some of the ways in which species differ:
a) Morphological differences (see Darwin’s definition, above). Morphology differs between races and populations, as well, of course; as already mentioned.
b) Enzyme and molecular differences. Francisco Ayala did detailed surveys with allozymes on Drosophila [SEE OVERHEAD]. Species differ at multiple allozyme loci, subspecies at slightly less loci, and so on down to poplations. We have seen that many hybrid zones separate subspecific forms that differ at multiple genetic loci also, and this and Ayala’s work shows clearly that races and species differ genetically in degree rather than kind. This is as true for mtDNA and other DNA markers as it is for allozymes (see also (g) below). Because multilocus differences are common even between populations and races that noone would want to call species, it is almost certain that speciation also involves multiple locus evolution, and indeed more of it!
c) Chromosomal differences. We have already mentioned human/chimp diffferences (see Chromosomal Evolution), and how common this is in other species that have been studied. Again we can point to subspecies and races that differ chromosomally also, only less so. Chromosomally, species are continuous with races, but usually differ more.
Polyploidy is, however one exception to this gradual differentiation. Polyploidy is a very common feature of plant species differences, and only rarely can be considered polymorphic within species because of the almost universal sterility of diploid X polyploid offspring, which are triploid.
d) Signals used in mating. Sexually-selected colours, tail length in birds, pheromones in moths, other insects, and even mammals are all involved in species recognition as well. In many crickets and grasshoppers, as well as frogs, species-specific sounds are required; in fireflies, species recognize each other by means of coded flashes [SEE OVERHEAD].
Again, these kinds differences are quite easily derived from mate choice differences within species, perhaps caused by sexual selection or for ecological reasons of efficiency. Differences between races and species are again in degree rather than kind. There is a controversy as to whether mate choice itself may evolve to "protect species" from gene flow. This would be a true isolating "mechanism". See next lecture.
e) Hybrid inviability and sterility - genomic incompatibility. Sterility and inviability are very common in hybrids. We have already mentioned examples produced by chromosomal differences. Mules (donkey x horse hybrids, which are sterile) are another example.
We know from studies of clines and hybrid zones that multilocus hybrid inviability can occur within species as well as between them. On the other hand, some species almost never mate together, but if they do, the hybrids seem not only viable but fertile. Related species of Darwin’s finches and ducks are an example. Once again, species differ from races only in the degree of hybrid inviability and sterility, not absolutely in kind.
A particularly well known kind of difference is known as Haldane's Rule after its discoverer, J.B.S. Haldane. Haldane's Rule states that when only one sex of the F1 hybrid between species is affected by inviability or sterility, that sex is usually the heterogametic (XY) sex, rather than the homogametic (XX) sex. The rule works in mammals and Diptera (flies) in which the sex-determination is usually male - XY, female - XX; as well as in birds and butterflies, in which females are XY and the males are XX. The reason is probably mainly due to recessive effects of genes causing incompatibility on the X chromosome. These genes must be epistatic; can you see why?
In other cases, the F1 hybrid between two species may be alright, but backcrosses or F2 crosses produce inviability or sterility. This is known as hybrid breakdown, and may be caused by recessive incompatibility genes (also epistatic) becoming homozygous during these later crosses.
f) Ecological differences. Perhaps the best examples we have of ecological differences between closely-related species are adaptive radiations on islands. Darwin’s finches are well-known. The Hawaiian honeycreepers [SEE OVERHEAD] are even more extraordinary. From finch-like ancestors, they have produced nectarivorous, insectivorous, frugivorous, as well as seed-eating forms.
But we have already discussed under the ecological species concept how ecological differences are found across clines that are under extrinsic selection across an environmental gradient. Once again there is no clear dividing line between races and species in the degree of ecological differentiation.
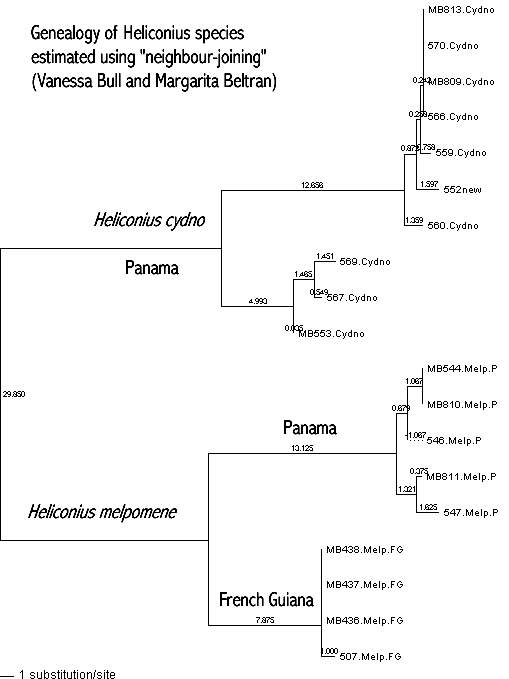
g) Genealogical differences. As we have seen, when species diverge, their DNA, such as mitochondrial DNA, will also diverge. When a genealogy (the phylogeny of a single gene or stretch of DNA) is estimated, one usually finds that species, and sometimes even races, fall on different branches of the genealogy. An example is given by the Heliconius butterflies, on which own group work, in the figure below.
Heliconius cydno and H. melpomene are closely related species which also occasionally hybridize. They clearly fall on separate branches of this genealogy of the genes CO1 and CO2 of mtDNA. However, it is also true that the melpomene from French Guiana falls on a separate branch of the genealogy from the members of the same species from Panama, and there is a similar deep branching pattern even within Panamanian cydno. Thus, a separate genealogy is not a good guide to separate species status. Other geographic races of Heliconius melpomene have mtDNA genealogies that intermingle with the Panamanian H. melpomene, so not all geographic populations have separate genealogies.
However, in some cases, as in the Drosophila (melanogaster, simulans, sechellia, and mauritiana) genealogies [OVERHEAD], gene genealogies of well-recognized separate species intermingle. In this case two possibilities exist: (1) ancestral polymorphisms -- speciation occurred recently enough so that polymorphisms for genes within each species are retained. (2) interspecific gene transfer -- horizontal gene transfer since the origin of the species has led to an intermingling of the genealogies more recently. These two are difficult to tell apart.
In any case, even at the genealogical level we see intermingling above the species level as well as below. Separate genealogical branches have evolved within some species, as well as between many, perhaps most species. Genealogies of species may be more separate than those of races and populations within species, but there is a lot of overlap.
Genetic differences
between species, then are usually inherited at multiple loci,
and are on average greater than and involve more genes than (though overlap
and blend into) the kinds of differences we see between geographic races,
or even morphs in polymorphic populations. There is nothing magic about
the species level in terms of genetics, and therefore it would seem most
logical and parsimonious (simplest) to use the same microevolutionary
forces - selection, drift, mutation - coupled with more time, to explain
the evoloution of species, as well as the other kinds of subspecific evolution
we have already discussed.
Strictly, biodiversity means the sum total of diversity at all levels of the evolutionary hierarchy, from genetic diversity within populations, between populations, between races, species, genera, and so on, up to ecosystems and biomes. In practice, the species is traditionally viewed as one of the most important level of biodiversity. In view of the difficulty of defining species (above), perhaps this isn't valid?
A good and also enjoyable summary of the
new science of biodiversity is in E.O. Wilson’s The diversity of life
(1992), from where I get figures for the numbers of species. An interesting
feature of the diversity of life is that most of it is NOT like us; it
is not mammalian, or even vertebrate or chordate. J.B.S. Haldane was once
asked what he thought that his studies in biology told him about the Mind
of the Creator. He replied "an inordinate fondness for beetles". [ See
overhead ]
| Total species described |
|
| Animals |
|
|
|
|
|
| Plants and fungi |
|
You can see that the humble beetle does make up a big proportion of life at the species level; about 20%. But other insects are also no slouchers, with insects making up about 53% of species. If you want to do important work in biodiversity be an entomologist!
Now many people complain about the bias
inherent in using species already described. Beetle systematists
have calculated, based on fogging the canopies of Amazonian tree species
for unidentified, host-specific beetles, that there may be as much as a
30-fold underestimate of animal species; 30,000,000 is their estimate.
Bacteriologists, in contrast, argue that the prokaryotic world is far more
diverse in terms, both of DNA diversity, and also probably in species,
than the eukaryotes. It is just that we haven’t discovered most of it yet.
Animals, plants, and fungi, the eukaryotes are now known to form only a
tiny sub-branch of the grand "tree of life", most of which is prokaryotic.
But the assertions about species numbers have yet to be proved.
FUTUYMA, DJ 1998.
Evolutionary Biology. Chapter 15 (pp. 447-479). Species.
WILSON, EO 1992.
The diversity of life.
Science Library:
View 2007 Teaching Collection by going to eUCLid;
use Keyword, Basic Search, All Fields: 2007
Next time:
how does all this diversity evolve?